

CXCR3 deficiency enhances tumor progression by promoting macrophage M2 polarization in a murine breast cancer model
- PMID: 24679047
- PMCID: PMC4137960
- DOI: 10.1111/imm.12293
CXCR3 deficiency enhances tumor progression by promoting macrophage M2 polarization in a murine breast cancer model
Abstract
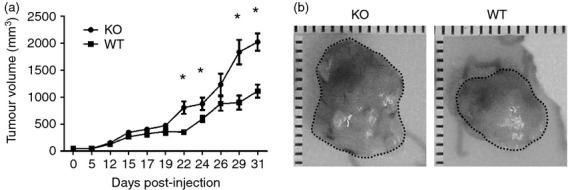
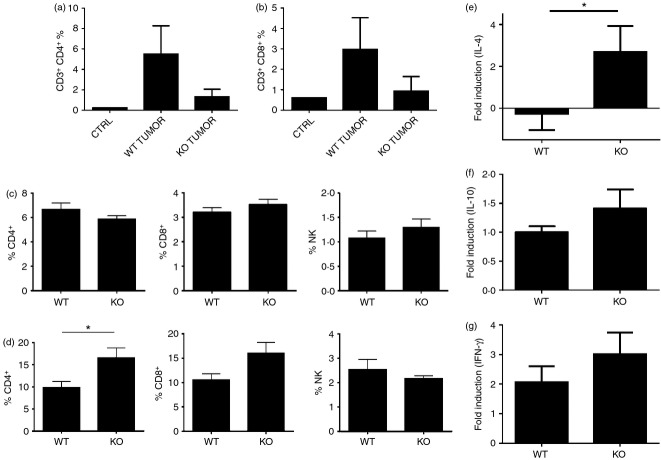
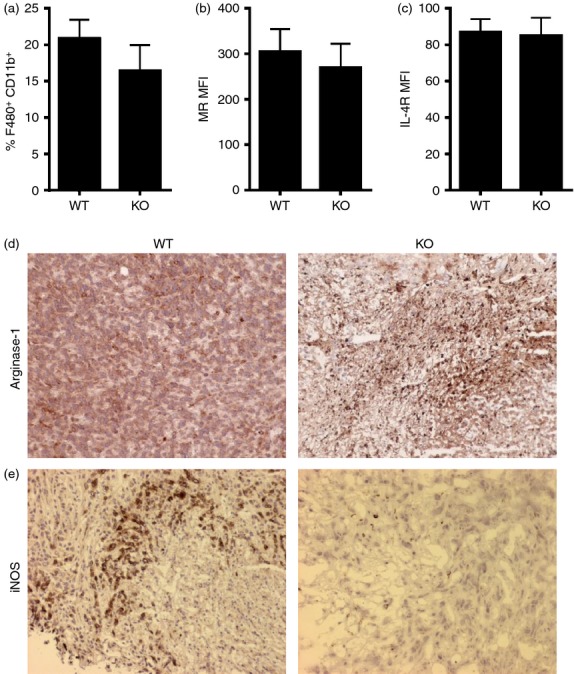
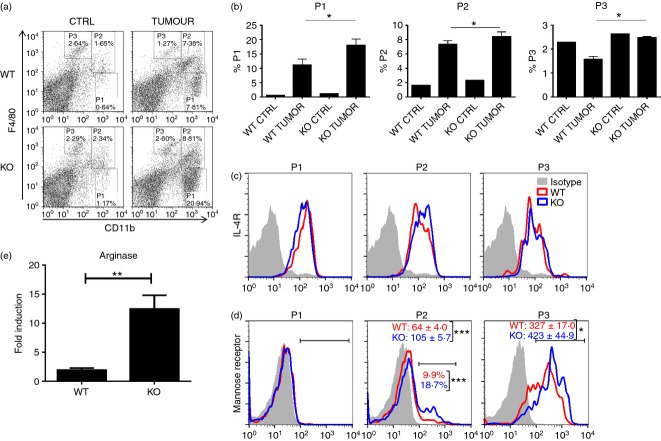
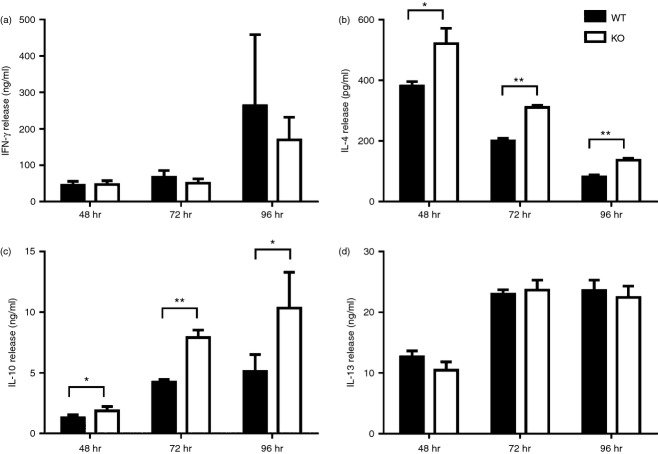
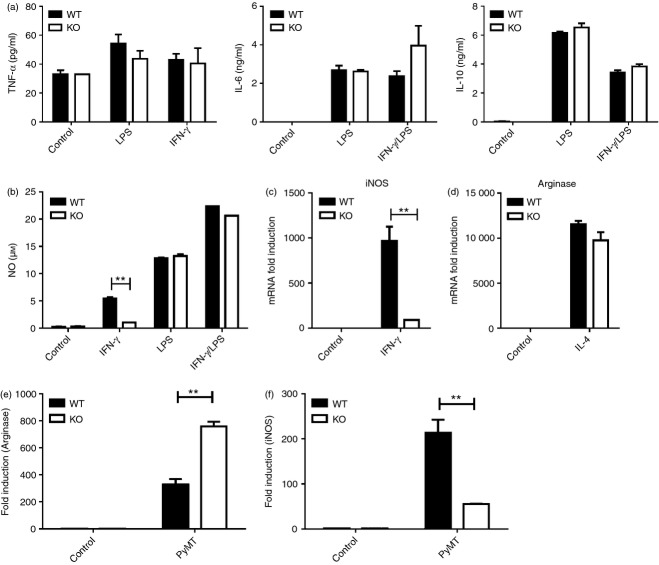
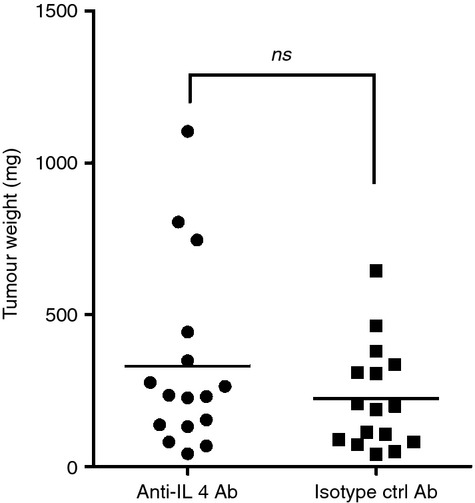
Tumor associated macrophages play a vital role in determining the outcome of breast cancer. We investigated the contribution of the chemokine receptor CXCR3 to antitumor immune responses using a cxcr3 deficient mouse orthotopically injected with a PyMT breast cancer cell line. We observed that cxcr3 deficient mice displayed increased IL-4 production and M2 polarization in the tumors and spleens compared to WT mice injected with PyMT cells. This was accompanied by larger tumor development in cxcr3(-/-) than in WT mice. Further, tumor-promoting myeloid derived immune cell populations accumulated in higher proportions in the spleens of cxcr3 deficient mice. Interestingly, cxcr3(-/-) macrophages displayed a deficiency in up-regulating inducible nitric oxide synthase after stimulation by either IFN-γ or PyMT supernatants. Stimulation of bone marrow derived macrophages by PyMT supernatants also resulted in greater induction of arginase-1 in cxcr3(-/-) than WT mice. Further, cxcr3(-/-) T cells activated with CD3/CD28 in vitro produced greater amounts of IL-4 and IL-10 than T cells from WT mice. Our data suggests that a greater predisposition of cxcr3 deficient macrophages towards M2 polarization contributes to an enhanced tumor promoting environment in cxcr3 deficient mice. Although CXCR3 is known to be expressed on some macrophages, this is the first report that demonstrates a role for CXCR3 in macrophage polarization and subsequent breast tumor outcomes. Targeting CXCR3 could be a potential therapeutic approach in the management of breast cancer tumors.
Keywords: CXCR3; arginase-1; breast cancer; inducible nitric oxide synthase; tumor associated macrophages.
© 2014 John Wiley & Sons Ltd.
Figures
References
-
- Galdiero MR, Garlanda C, Jaillon S, Marone G, Mantovani A. Tumor associated macrophages and neutrophils in tumor progression. J Cell Physiol. 2013;228:1404–12. - PubMed
-
- Menard S, Tomasic G, Casalini P, et al. Lymphoid infiltration as a prognostic variable for early-onset breast carcinomas. Clin Cancer Res. 1997;3:817–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
