

To be or not to be? How selective autophagy and cell death govern cell fate
- PMID: 24679527
- PMCID: PMC4020175
- DOI: 10.1016/j.cell.2014.02.049
To be or not to be? How selective autophagy and cell death govern cell fate
Abstract
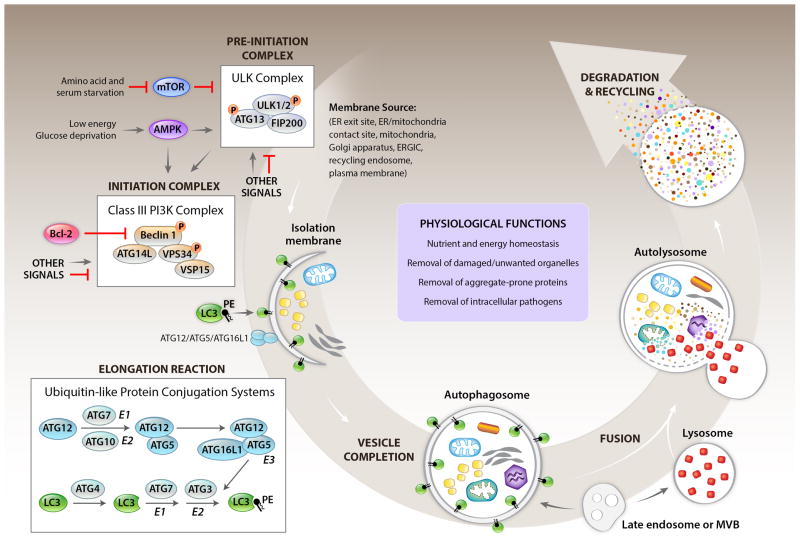
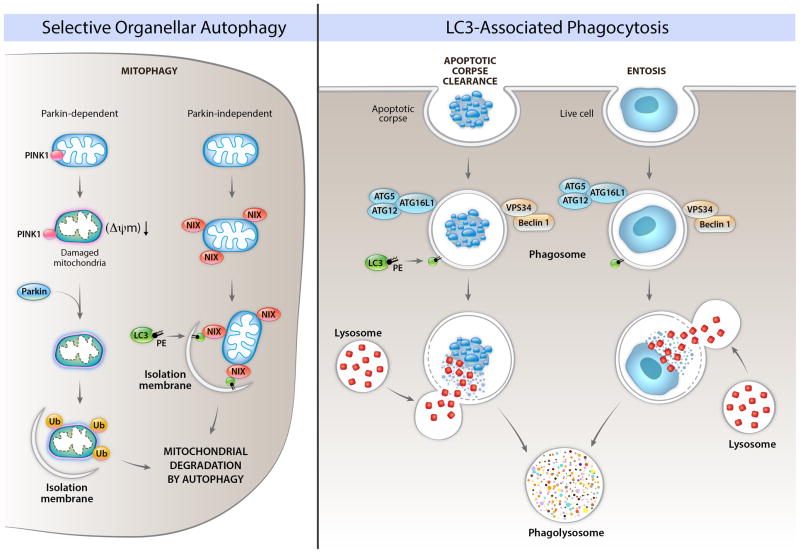
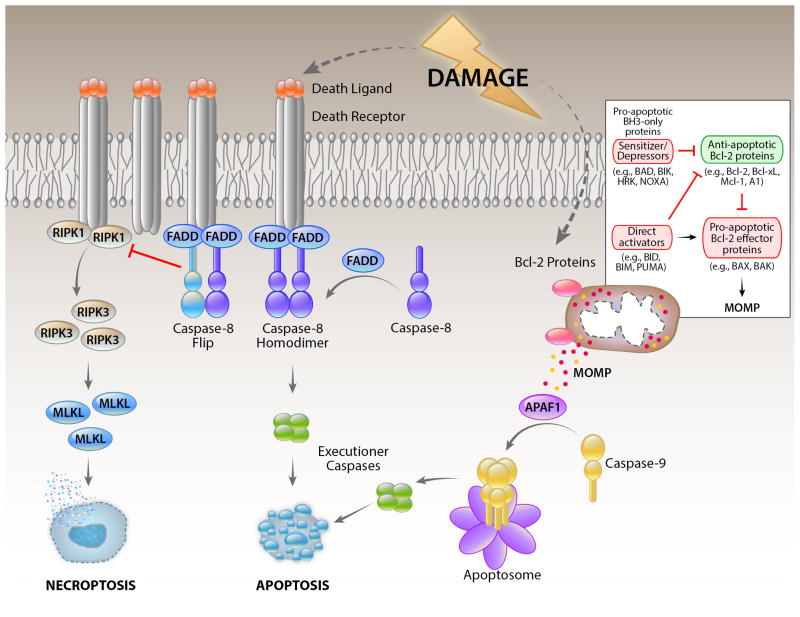
The health of metazoan organisms requires an effective response to organellar and cellular damage either by repair of such damage and/or by elimination of the damaged parts of the cells or the damaged cell in its entirety. Here, we consider the progress that has been made in the last few decades in determining the fates of damaged organelles and damaged cells through discrete, but genetically overlapping, pathways involving the selective autophagy and cell death machinery. We further discuss the ways in which the autophagy machinery may impact the clearance and consequences of dying cells for host physiology. Failure in the proper removal of damaged organelles and/or damaged cells by selective autophagy and cell death processes is likely to contribute to developmental abnormalities, cancer, aging, inflammation, and other diseases.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Al Rawi S, Louvet-Vallee S, Djeddi A, Sachse M, Culetto E, Hajjar C, Boyd L, Legouis R, Galy V. Postfertilization autophagy of sperm organelles prevents paternal mitochondrial DNA transmission. Science. 2011;334:1144–1147. - PubMed
-
- Chao MP, Majeti R, Weissman IL. Programmed cell removal: a new obstacle in the road to developing cancer. Nat Rev Cancer. 2011;12:58–67. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
