

Monitoring Keap1-Nrf2 interactions in single live cells
- PMID: 24681086
- PMCID: PMC4165437
- DOI: 10.1016/j.biotechadv.2014.03.004
Monitoring Keap1-Nrf2 interactions in single live cells
Abstract
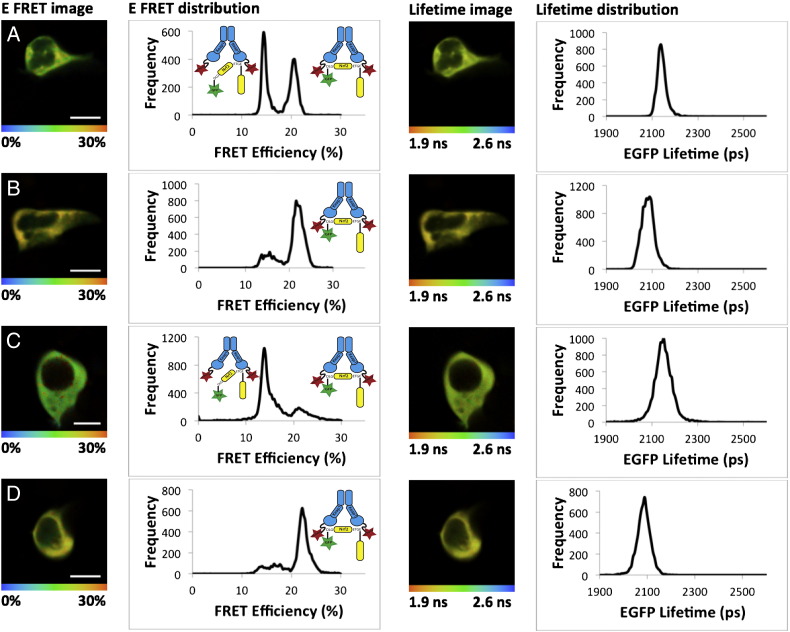
The transcription factor NF-E2 p45-related factor 2 (Nrf2) and its negative regulator Kelch-like ECH associated protein 1 (Keap1) control the expression of nearly 500 genes with diverse cytoprotective functions. Keap1, a substrate adaptor protein for Cullin3/Rbx1 ubiquitin ligase, normally continuously targets Nrf2 for degradation, but loses this ability in response to electrophiles and oxidants (termed inducers). Consequently, Nrf2 accumulates and activates transcription of its downstream target genes. Many inducers are phytochemicals, and cruciferous vegetables represent one of the richest sources of inducer activity among the most commonly used edible plants. Here we summarize the discovery of the isothiocyanate sulforaphane as a potent inducer which reacts with cysteine sensors of Keap1, leading to activation of Nrf2. We then describe the development of a quantitative Förster resonance energy transfer (FRET)-based methodology combined with multiphoton fluorescence lifetime imaging microscopy (FLIM) to investigate the interactions between Keap1 and Nrf2 in single live cells, and the effect of sulforaphane, and other cysteine-reactive inducers, on the dynamics of the Keap1-Nrf2 protein complex. We present the experimental evidence for the "cyclic sequential attachment and regeneration" or "conformation cycling" model of Keap1-mediated Nrf2 degradation. Finally, we discuss the implications of this mode of regulation of Nrf2 for achieving a fine balance under normal physiological conditions, and the consequences and mechanisms of disrupting this balance for tumor biology.
Keywords: Cytoprotective enzymes; FLIM; FRET; Keap1; Nrf2; Sulforaphane.
Copyright © 2014. Published by Elsevier Inc.
Figures





References
-
- Bae S.H., Sung S.H., Oh S.Y., Lim J.M., Lee S.K., Park Y.N., et al. Sestrins activate Nrf2 by promoting p62-dependent autophagic degradation of Keap1 and prevent oxidative liver damage. Cell Metab. 2013;17:73–84. - PubMed
-
- Baird L., Dinkova-Kostova A.T. The cytoprotective role of the Keap1–Nrf2 pathway. Arch Toxicol. 2011;85:241–272. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
