

Resistance of Biomphalaria glabrata 13-16-R1 snails to Schistosoma mansoni PR1 is a function of haemocyte abundance and constitutive levels of specific transcripts in haemocytes
- PMID: 24681237
- PMCID: PMC4096035
- DOI: 10.1016/j.ijpara.2013.11.004
Resistance of Biomphalaria glabrata 13-16-R1 snails to Schistosoma mansoni PR1 is a function of haemocyte abundance and constitutive levels of specific transcripts in haemocytes
Abstract
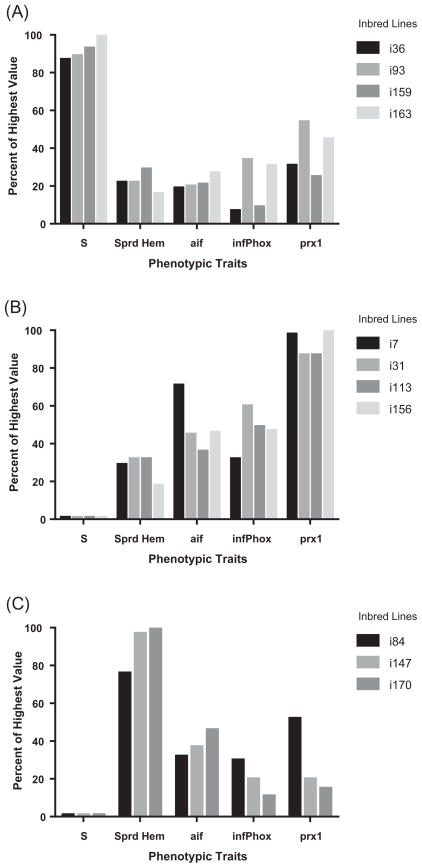
Continuing transmission of human intestinal schistosomiasis depends on the parasite's access to susceptible snail intermediate hosts (often Biomphalaria glabrata). Transmission fails when parasite larvae enter resistant individuals in wild snail populations. The genetic basis for differences in snail susceptibility/resistance is being intensively investigated as a means to devise novel control strategies based on resistance genes. Reactive oxygen species produced by the snail's defence cells (haemocytes) are effectors of resistance. We hypothesised that genes relevant to production and consumption of reactive oxygen species would be expressed differentially in the haemocytes of snail hosts with different susceptibility/resistance phenotypes. By restricting the genetic diversity of snails, we sought to facilitate identification of resistance genes. By inbreeding, we procured from a 13-16-R1 snail population with both susceptible and resistant individuals 52 lines of B. glabrata (expected homozygosity ~87.5%), and determined the phenotype of each in regard to susceptibility/resistance to Schistosoma mansoni. The inbred lines were found to have line-specific differences in numbers of spreading haemocytes; these were enumerated in both juvenile and adult snails. Lines with high cell numbers were invariably resistant to S. mansoni, whereas lines with lower cell numbers could be resistant or susceptible. Transcript levels in haemocytes were quantified for 18 potentially defence-related genes. Among snails with low cell numbers, the different susceptibility/resistance phenotypes correlated with differences in transcript levels for two redox-relevant genes: an inferred phagocyte oxidase component and a peroxiredoxin. Allograft inflammatory factor (potentially a regulator of leucocyte activation) was expressed at higher levels in resistant snails regardless of spread cell number. Having abundant spreading haemocytes is inferred to enable a snail to kill parasite sporocysts. In contrast, snails with fewer spreading haemocytes seem to achieve resistance only if specific genes are expressed constitutively at levels that are high for the species.
Keywords: Biomphalaria; Expression; Haematocrit; Haemocyte; Host–parasite; Oxidative; Resistance; Schistosoma.
Copyright © 2014 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Figures




References
-
- Adema CM, Hanington PC, Lun CM, Rosenberg GH, Aragon AD, Stout BA, Lennard Richard ML, Gross PS, Loker ES. Differential transcriptomic responses of Biomphalaria glabrata (Gastropoda, Mollusca) to bacteria and metazoan parasites, Schistosoma mansoni and Echinostoma paraensei (Digenea, Platyhelminthes) Mol Immunol. 2010;47:849–860. http://dx.doi.org/10.1016/j.molimm.2009.10.019. - DOI - PMC - PubMed
-
- Baeza Garcia A, Pierce RJ, Gourbal B, Werkmeister E, Colinet D, Reichhart JM, Dissous C, Coustau C. Involvement of the cytokine MIF in the snail host immune response to the parasite Schistosoma mansoni. PLoS Pathog. 2010;6 (9):e1001115. http://dx.doi.org/10.1371/journal.ppat.1001115. - DOI - PMC - PubMed
-
- Barçante TA, Barçante JM, Fujiwara RT, Lima WS. Analysis of circulating haemocytes from Biomphalaria glabrata following Angiostrongylus vasorum infection using flow cytometry. J Parasitol Res. 2012;2012:314723. http://dx.doi.org/10.1155/2012/314723. - DOI - PMC - PubMed
-
- Basch PF. An interpretation of snail-trematode infection rates: specificity based on concordance of compatible phenotypes. Int J Parasitol. 1975;5:449–452. - PubMed
-
- Bayne CJ. Successful parasitism of vector snail Biomphalaria glabrata by the human blood fluke (trematode) Schistosoma mansoni: a 2009 assessment. Mol Biochem Parasitol. 2009;165:8–18. http://dx.doi.org/10.1016/j.molbiopara.2009.01.005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
