

The past, present, and future of human centromere genomics
- PMID: 24683489
- PMCID: PMC3966626
- DOI: 10.3390/genes5010033
The past, present, and future of human centromere genomics
Abstract
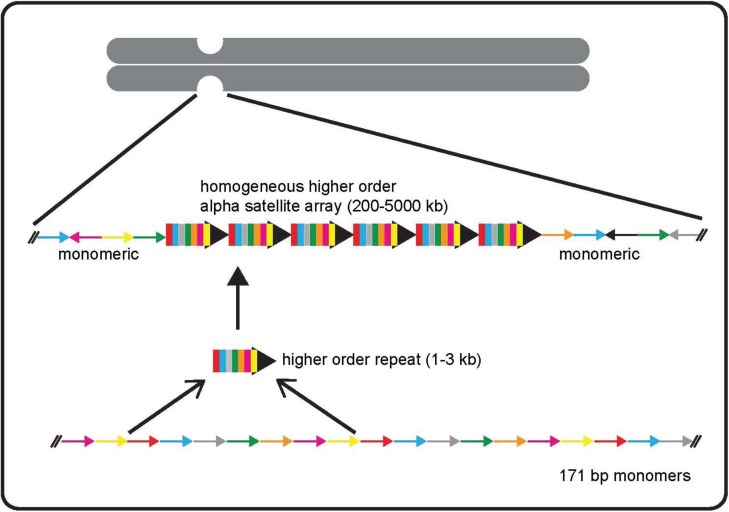
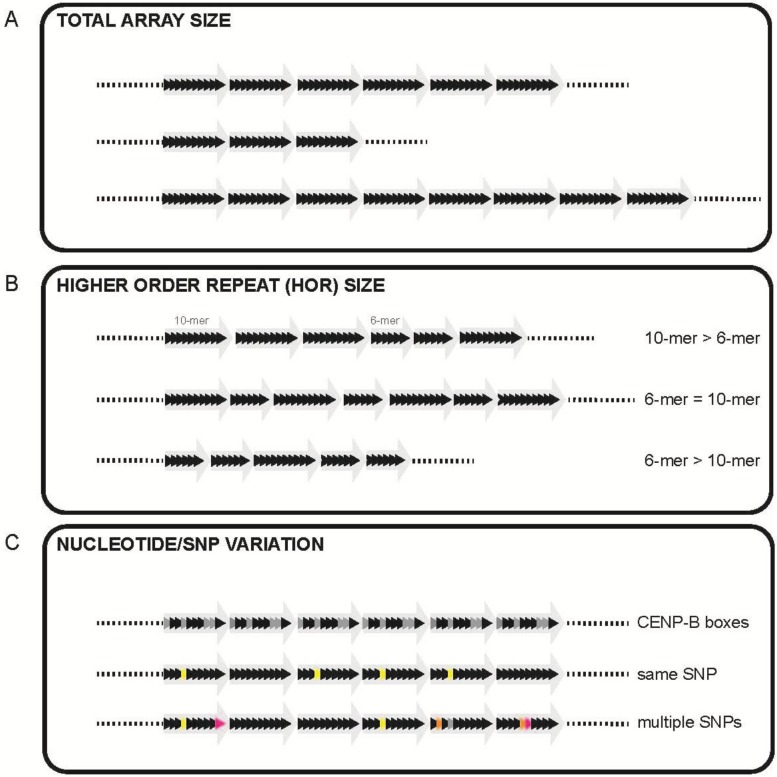
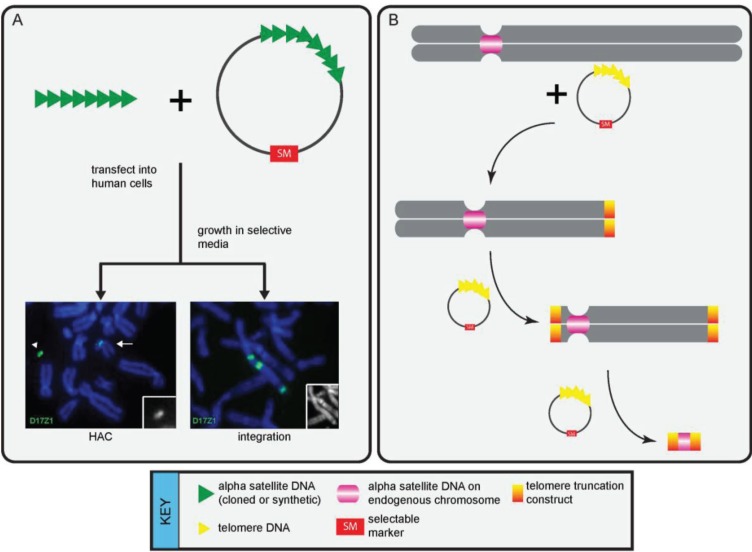
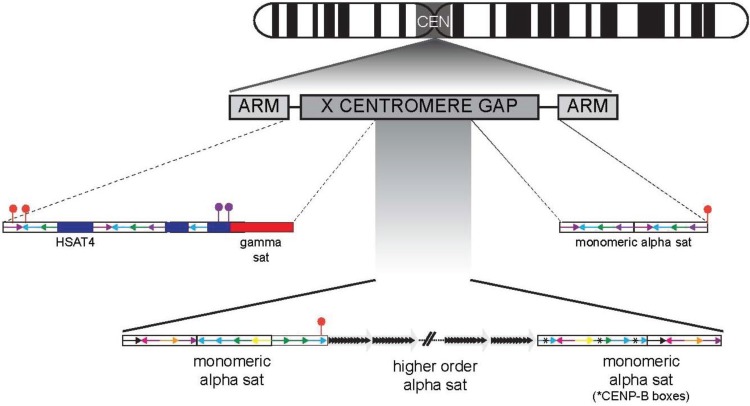
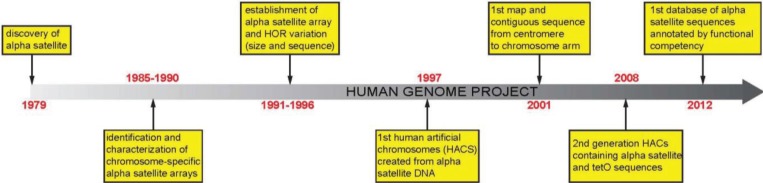
The centromere is the chromosomal locus essential for chromosome inheritance and genome stability. Human centromeres are located at repetitive alpha satellite DNA arrays that compose approximately 5% of the genome. Contiguous alpha satellite DNA sequence is absent from the assembled reference genome, limiting current understanding of centromere organization and function. Here, we review the progress in centromere genomics spanning the discovery of the sequence to its molecular characterization and the work done during the Human Genome Project era to elucidate alpha satellite structure and sequence variation. We discuss exciting recent advances in alpha satellite sequence assembly that have provided important insight into the abundance and complex organization of this sequence on human chromosomes. In light of these new findings, we offer perspectives for future studies of human centromere assembly and function.
Keywords: CENP; alpha satellite; chromosome truncation; dicentric; heterochromatin; higher order repeat; human artificial chromosome; tet operon; transcription.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous