

Arabinogalactan protein 31 (AGP31), a putative network-forming protein in Arabidopsis thaliana cell walls?
- PMID: 24685714
- PMCID: PMC4195544
- DOI: 10.1093/aob/mcu038
Arabinogalactan protein 31 (AGP31), a putative network-forming protein in Arabidopsis thaliana cell walls?
Abstract
Background and aims: Arabinogalactan protein 31 (AGP31) is a remarkable plant cell-wall protein displaying a multi-domain organization unique in Arabidopsis thaliana: it comprises a predicted signal peptide (SP), a short AGP domain of seven amino acids, a His-stretch, a Pro-rich domain and a PAC (PRP-AGP containing Cys) domain. AGP31 displays different O-glycosylation patterns with arabinogalactans on the AGP domain and Hyp-O-Gal/Ara-rich motifs on the Pro-rich domain. AGP31 has been identified as an abundant protein in cell walls of etiolated hypocotyls, but its function has not been investigated thus far. Literature data suggest that AGP31 may interact with cell-wall components. The purpose of the present study was to identify AGP31 partners to gain new insight into its function in cell walls.
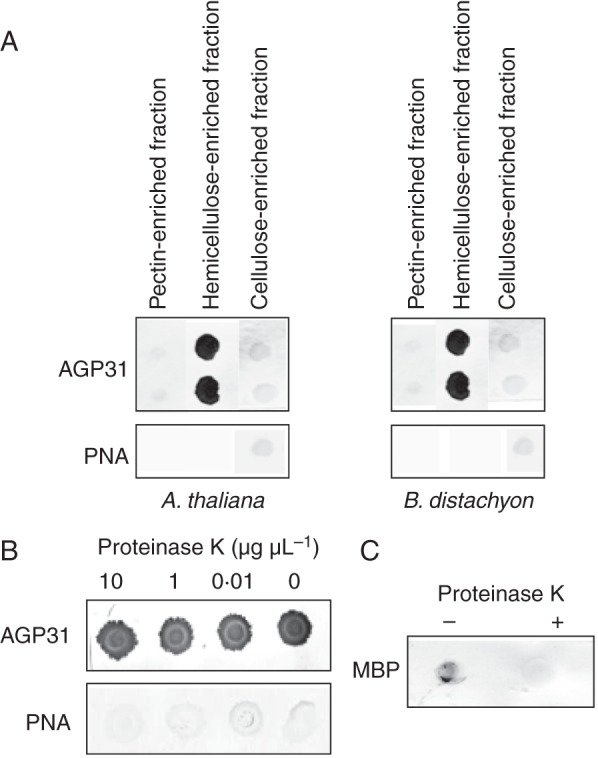
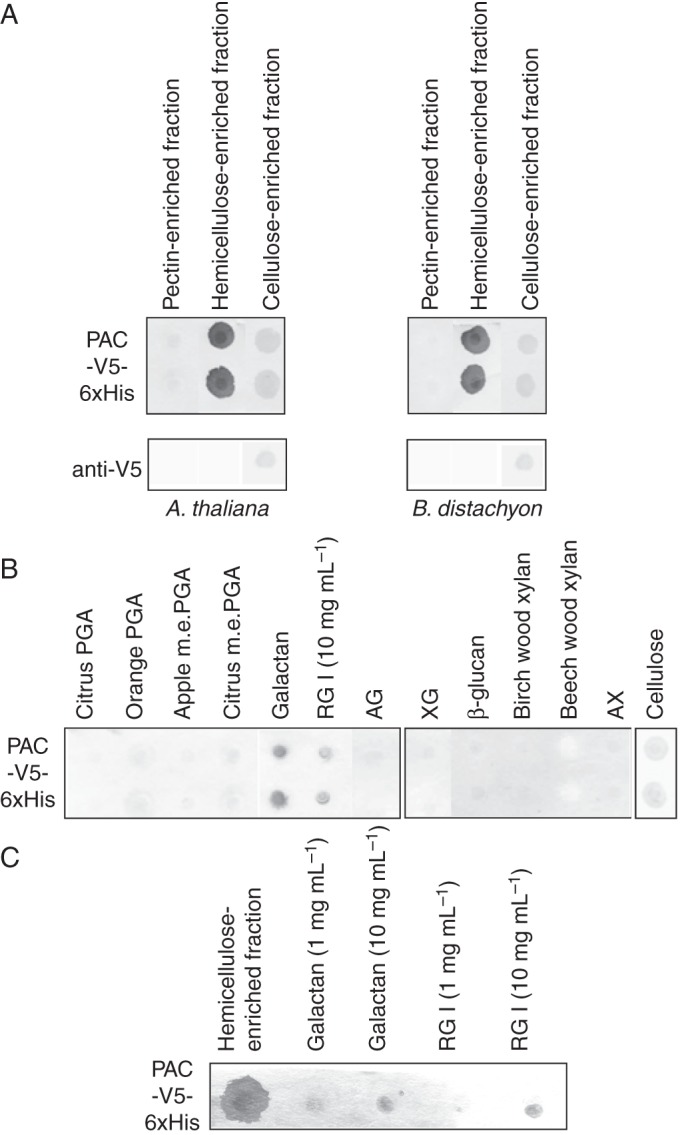
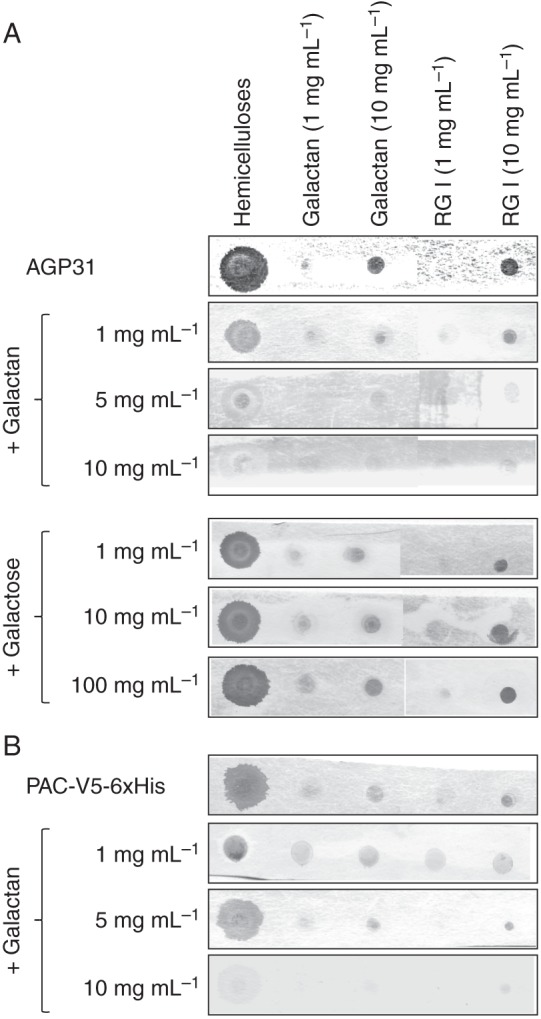
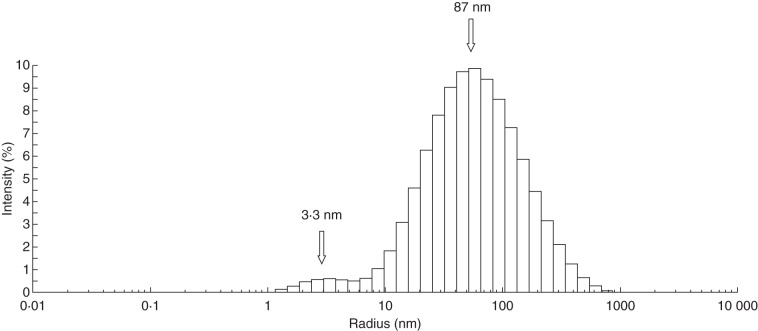
Methods: Nitrocellulose membranes were prepared by spotting different polysaccharides, which were either obtained commercially or extracted from cell walls of Arabidopsis thaliana and Brachypodium distachyon. After validation of the arrays, in vitro interaction assays were carried out by probing the membranes with purified native AGP31 or recombinant PAC-V5-6xHis. In addition, dynamic light scattering (DLS) analyses were carried out on an AGP31 purified fraction.
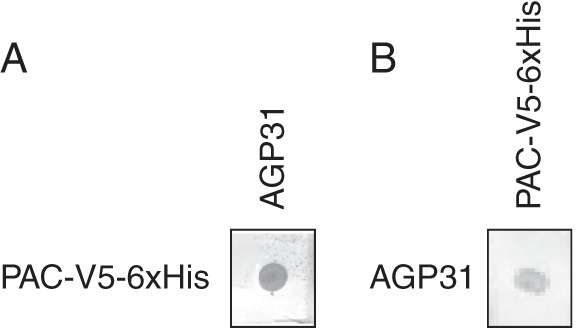
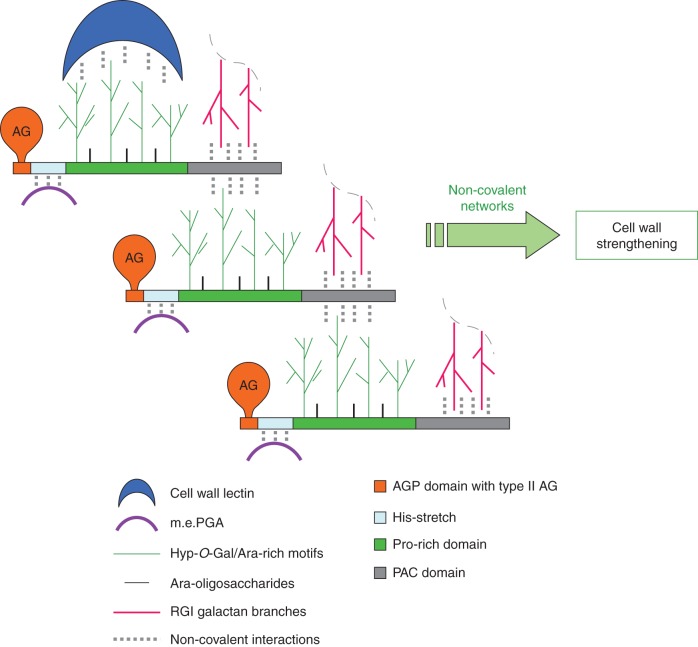
Key results: It was demonstrated that AGP31 interacts through its PAC domain with galactans that are branches of rhamnogalacturonan I. This is the first experimental evidence that a PAC domain, also found as an entire protein or a domain of AGP31 homologues, can bind carbohydrates. AGP31 was also found to bind methylesterified polygalacturonic acid, possibly through its His-stretch. Finally, AGP31 was able to interact with itself in vitro through its PAC domain. DLS data showed that AGP31 forms aggregates in solution, corroborating the hypothesis of an auto-assembly.
Conclusions: These results allow the proposal of a model of interactions of AGP31 with different cell-wall components, in which AGP31 participates in complex supra-molecular scaffolds. Such scaffolds could contribute to the strengthening of cell walls of quickly growing organs such as etiolated hypocotyls.
Keywords: AGP31; Arabidopsis thaliana; Brachypodium distachyon; PAC domain; arabinogalactan protein 31; glycoprotein; non-covalent interactions; plant cell wall; polysaccharides.
© The Author 2014. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures


Similar articles
-
Characterization of the arabinogalactan protein 31 (AGP31) of Arabidopsis thaliana: new advances on the Hyp-O-glycosylation of the Pro-rich domain.J Biol Chem. 2012 Mar 16;287(12):9623-32. doi: 10.1074/jbc.M111.247874. Epub 2012 Jan 23. J Biol Chem. 2012. PMID: 22270363 Free PMC article.
-
A nonclassical arabinogalactan protein gene highly expressed in vascular tissues, AGP31, is transcriptionally repressed by methyl jasmonic acid in Arabidopsis.Plant Physiol. 2007 Nov;145(3):863-74. doi: 10.1104/pp.107.102657. Epub 2007 Sep 20. Plant Physiol. 2007. PMID: 17885091 Free PMC article.
-
An Arabidopsis cell wall proteoglycan consists of pectin and arabinoxylan covalently linked to an arabinogalactan protein.Plant Cell. 2013 Jan;25(1):270-87. doi: 10.1105/tpc.112.107334. Epub 2013 Jan 31. Plant Cell. 2013. PMID: 23371948 Free PMC article.
-
Back to the future with the AGP-Ca2+ flux capacitor.Ann Bot. 2014 Oct;114(6):1069-85. doi: 10.1093/aob/mcu161. Epub 2014 Aug 19. Ann Bot. 2014. PMID: 25139429 Free PMC article. Review.
-
Plant Cell Wall Proteomics: A Focus on Monocot Species, Brachypodium distachyon, Saccharum spp. and Oryza sativa.Int J Mol Sci. 2019 Apr 23;20(8):1975. doi: 10.3390/ijms20081975. Int J Mol Sci. 2019. PMID: 31018495 Free PMC article. Review.
Cited by
-
Co-operative intermolecular kinetics of 2-oxoglutarate dependent dioxygenases may be essential for system-level regulation of plant cell physiology.Front Plant Sci. 2015 Jul 15;6:489. doi: 10.3389/fpls.2015.00489. eCollection 2015. Front Plant Sci. 2015. PMID: 26236316 Free PMC article.
-
The unique evolutionary pattern of the Hydroxyproline-rich glycoproteins superfamily in Chinese white pear (Pyrus bretschneideri).BMC Plant Biol. 2018 Feb 17;18(1):36. doi: 10.1186/s12870-018-1252-2. BMC Plant Biol. 2018. PMID: 29454308 Free PMC article.
-
An update on post-translational modifications of hydroxyproline-rich glycoproteins: toward a model highlighting their contribution to plant cell wall architecture.Front Plant Sci. 2014 Aug 14;5:395. doi: 10.3389/fpls.2014.00395. eCollection 2014. Front Plant Sci. 2014. PMID: 25177325 Free PMC article. Review.
-
Extensins: Self-Assembly, Crosslinking, and the Role of Peroxidases.Front Plant Sci. 2021 May 14;12:664738. doi: 10.3389/fpls.2021.664738. eCollection 2021. Front Plant Sci. 2021. PMID: 34054905 Free PMC article. Review.
-
Plant and algal cell walls: diversity and functionality.Ann Bot. 2014 Oct;114(6):1043-8. doi: 10.1093/aob/mcu214. Ann Bot. 2014. PMID: 25453142 Free PMC article.
References
-
- Baldwin T, Domingo C, Schindler T, Seetharaman G, Stacey N, Roberts K. DcAGP1, a secreted arabinogalactan protein, is related to a family of basic proline-rich proteins. Plant Molecular Biology. 2001;45:421–435. - PubMed
-
- Bradford MM. Rapid and sensitive method for quantification of microgram quantities of protein utilizing principle of protein-dye binding. Annals of Biochemistry. 1976;72:248–254. - PubMed
-
- Bradley DJ, Kjellbom P, Lamb CJ. Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: a novel, rapid defense response. Cell. 1992;70:21–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
