

Phase variation mediates reductions in expression of surface proteins during persistent meningococcal carriage
- PMID: 24686058
- PMCID: PMC4019173
- DOI: 10.1128/IAI.01521-14
Phase variation mediates reductions in expression of surface proteins during persistent meningococcal carriage
Abstract
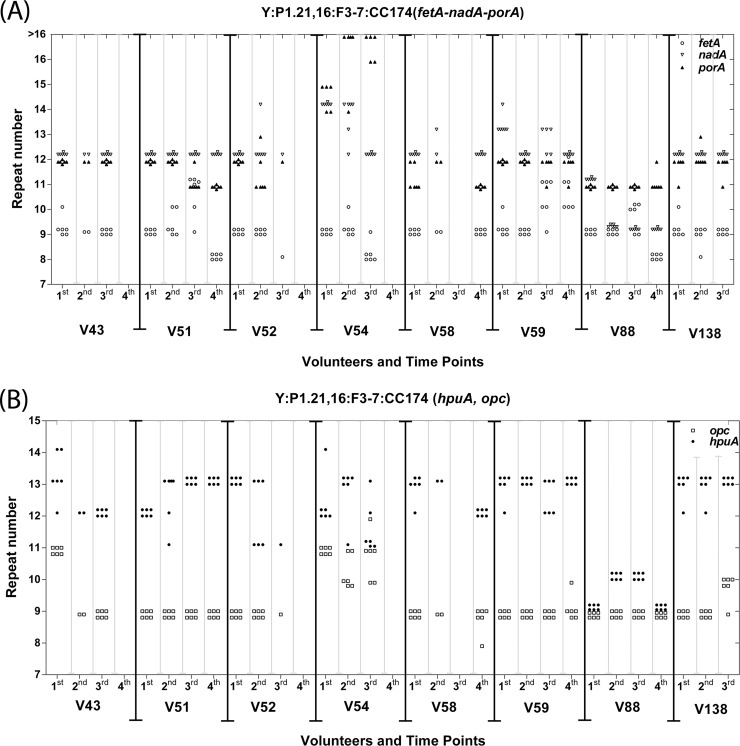
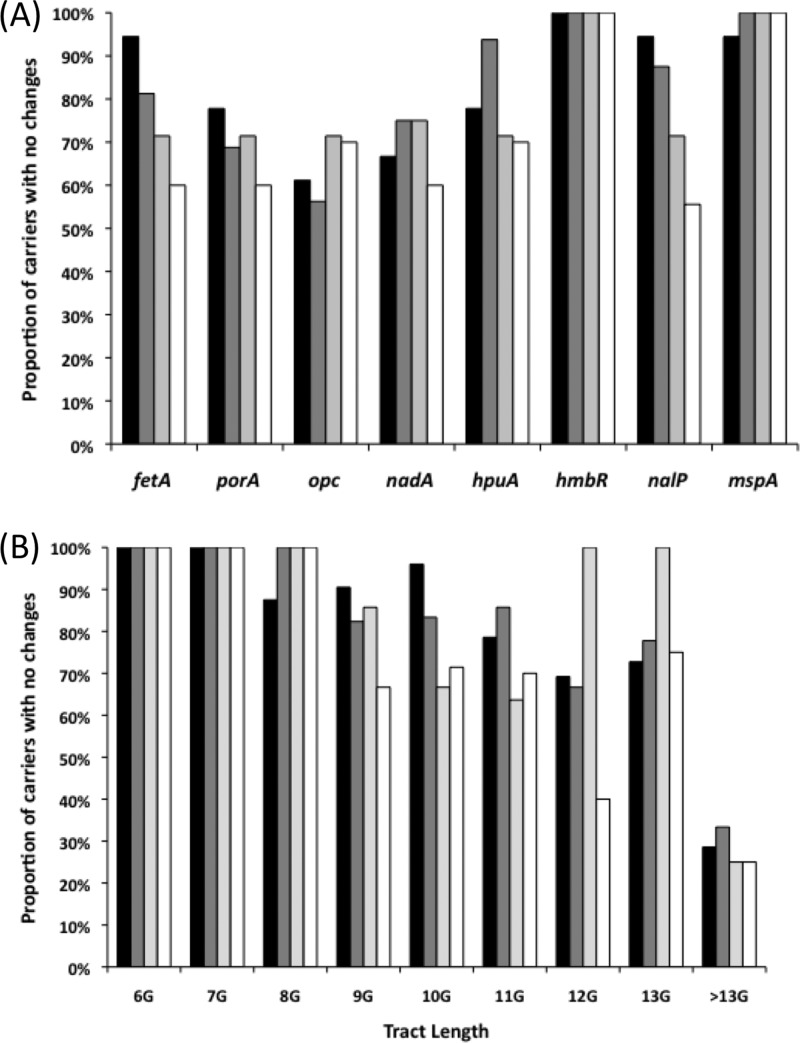
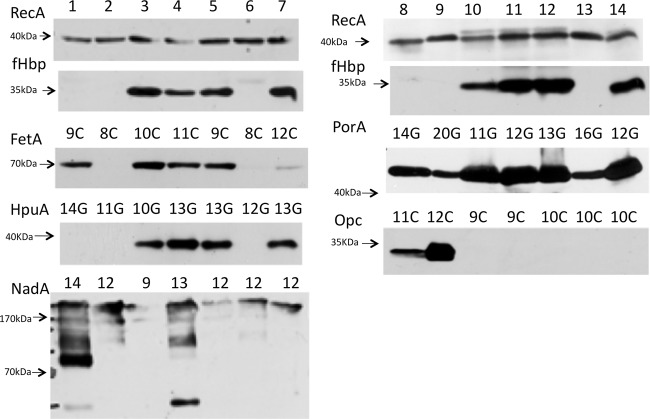
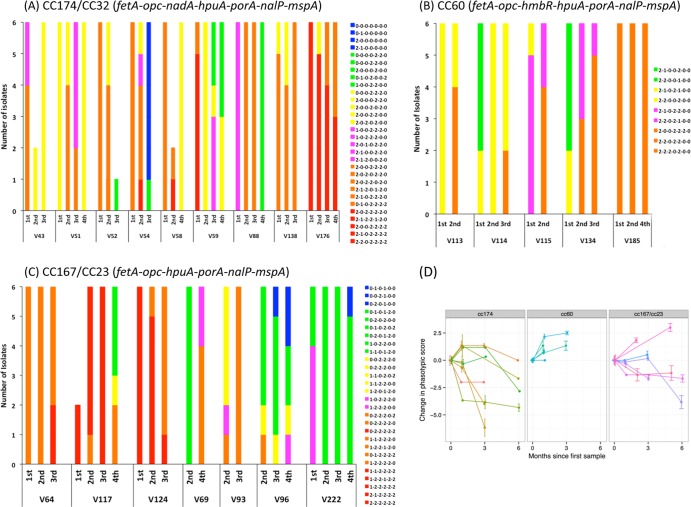
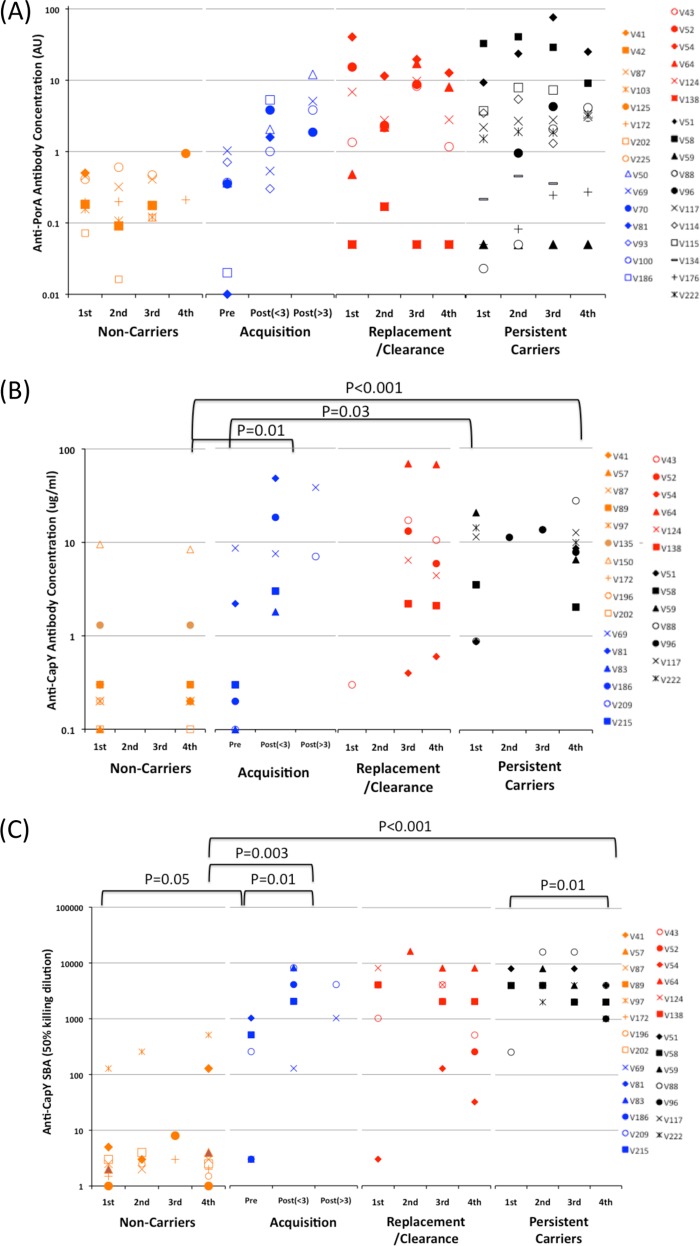
Asymptomatic and persistent colonization of the upper respiratory tract by Neisseria meningitidis occurs despite elicitation of adaptive immune responses against surface antigens. A putative mechanism for facilitating host persistence of this bacterial commensal and pathogen is alterations in expression of surface antigens by simple sequence repeat (SSR)-mediated phase variation. We investigated how often phase variation occurs during persistent carriage by analyzing the SSRs of eight loci in multiple isolates from 21 carriers representative of 1 to 6 months carriage. Alterations in repeat number were detected by a GeneScan analysis and occurred at 0.06 mutations/gene/month of carriage. The expression states were determined by Western blotting and two genes, fetA and nadA, exhibited trends toward low expression states. A critical finding from our unique examination of combinatorial expression states, "phasotypes," was for significant reductions in expression of multiple phase-variable surface proteins during persistent carriage of some strains. The immune responses in these carriers were examined by measuring variant-specific PorA IgG antibodies, capsular group Y IgG antibodies and serum bactericidal activity in concomitant serum samples. Persistent carriage was associated with high levels of specific IgG antibodies and serum bactericidal activity while recent strain acquisition correlated with a significant induction of antibodies. We conclude that phase-variable genes are driven into lower expression states during long-term persistent meningococcal carriage, in part due to continuous exposure to antibody-mediated selection, suggesting localized hypermutation has evolved to facilitate host persistence.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
