

Whisker barrel cortex delta oscillations and gamma power in the awake mouse are linked to respiration
- PMID: 24686563
- PMCID: PMC3988824
- DOI: 10.1038/ncomms4572
Whisker barrel cortex delta oscillations and gamma power in the awake mouse are linked to respiration
Abstract
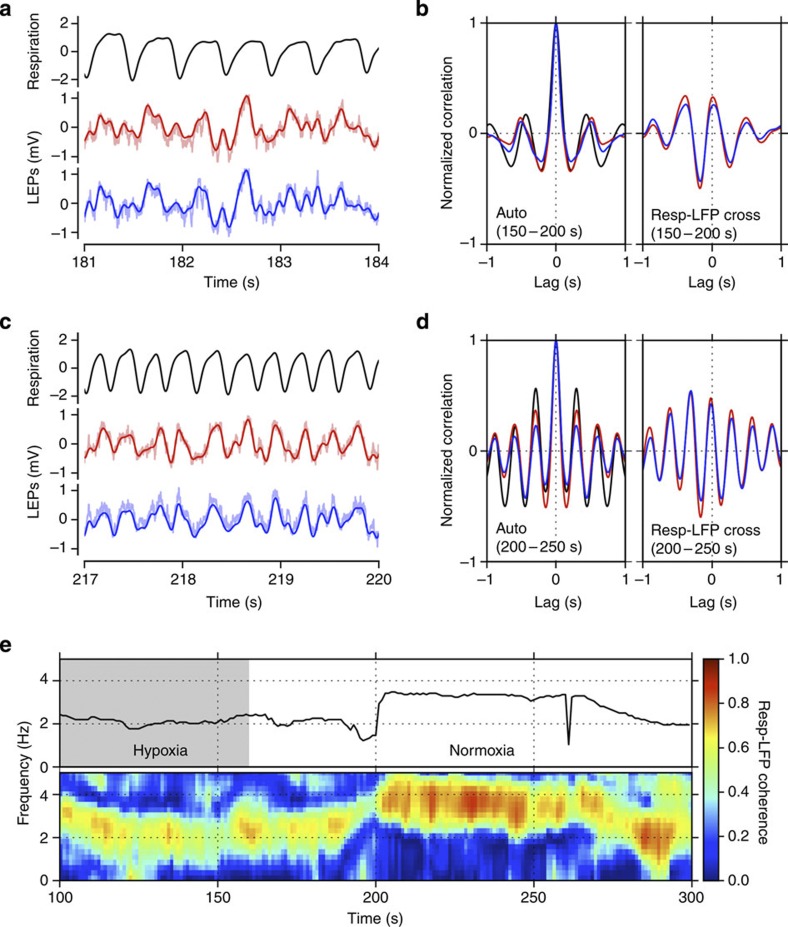
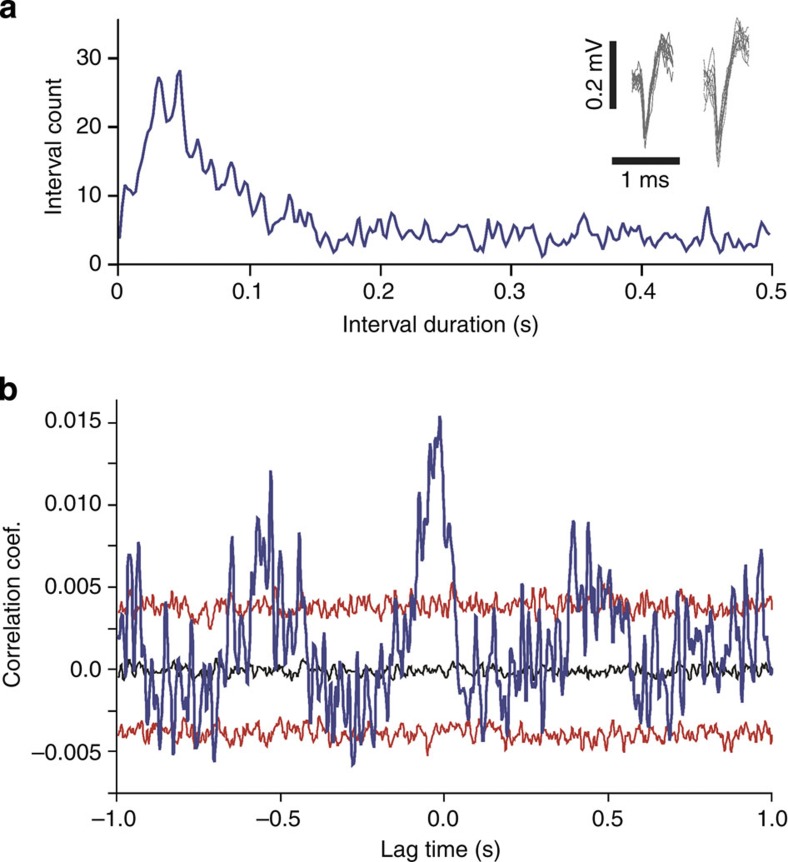
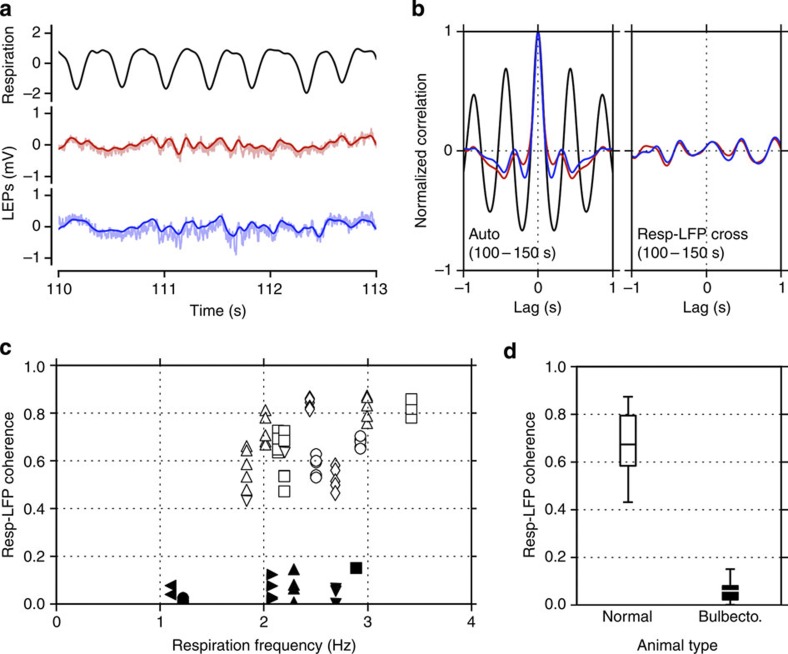
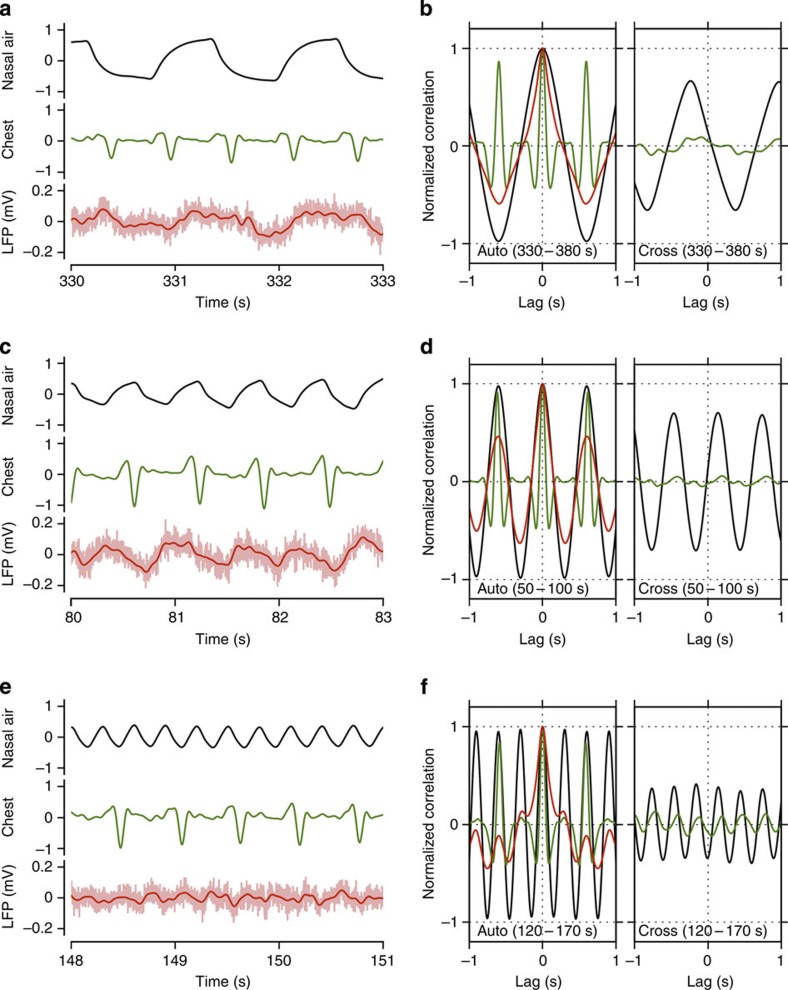
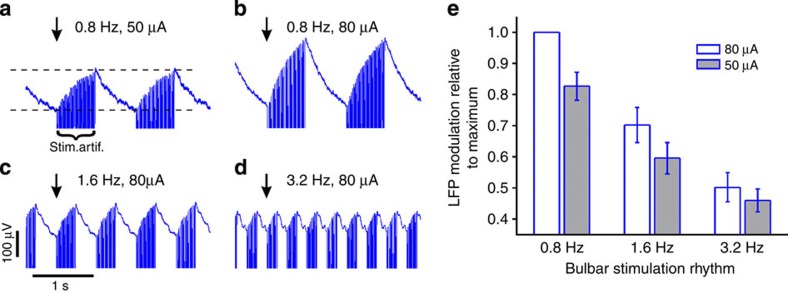
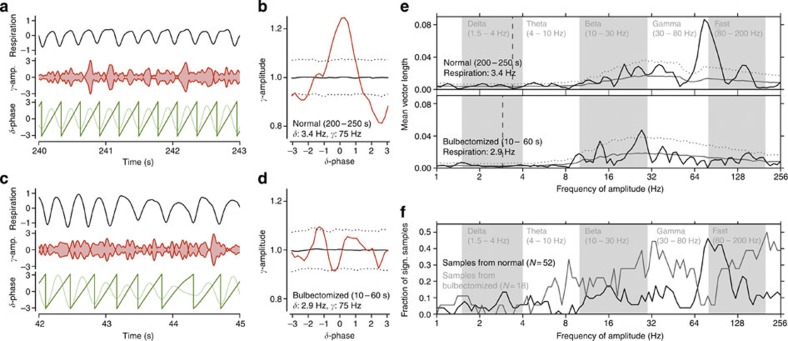
Current evidence suggests that delta oscillations (0.5-4 Hz) in the brain are generated by intrinsic network mechanisms involving cortical and thalamic circuits. Here we report that delta band oscillation in spike and local field potential (LFP) activity in the whisker barrel cortex of awake mice is phase locked to respiration. Furthermore, LFP oscillations in the gamma frequency band (30-80 Hz) are amplitude modulated in phase with the respiratory rhythm. Removal of the olfactory bulb eliminates respiration-locked delta oscillations and delta-gamma phase-amplitude coupling. Our findings thus suggest respiration-locked olfactory bulb activity as a main driving force behind delta oscillations and gamma power modulation in the whisker barrel cortex in the awake state.
Figures

References
-
- Buzsaki G. Rhythms of the Brain Oxford University Press (2006).
-
- Buzsaki G. & Draguhn A. Neuronal oscillations in cortical networks. Science 304, 1926–1929 (2004). - PubMed
-
- Amzica F. & Steriade M. Electrophysiological correlates of sleep delta waves. Electroencephalogr. Clin. Neurophysiol. 107, 69–83 (1998). - PubMed
-
- Timofeev I. & Chauvette S. Thalamocortical oscillations: local control of EEG slow waves. Curr. Top. Med. Chem. 11, 2457–2471 (2011). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
