

Genetic conflict, kin and the origins of novel genetic systems
- PMID: 24686935
- PMCID: PMC3982665
- DOI: 10.1098/rstb.2013.0364
Genetic conflict, kin and the origins of novel genetic systems
Abstract
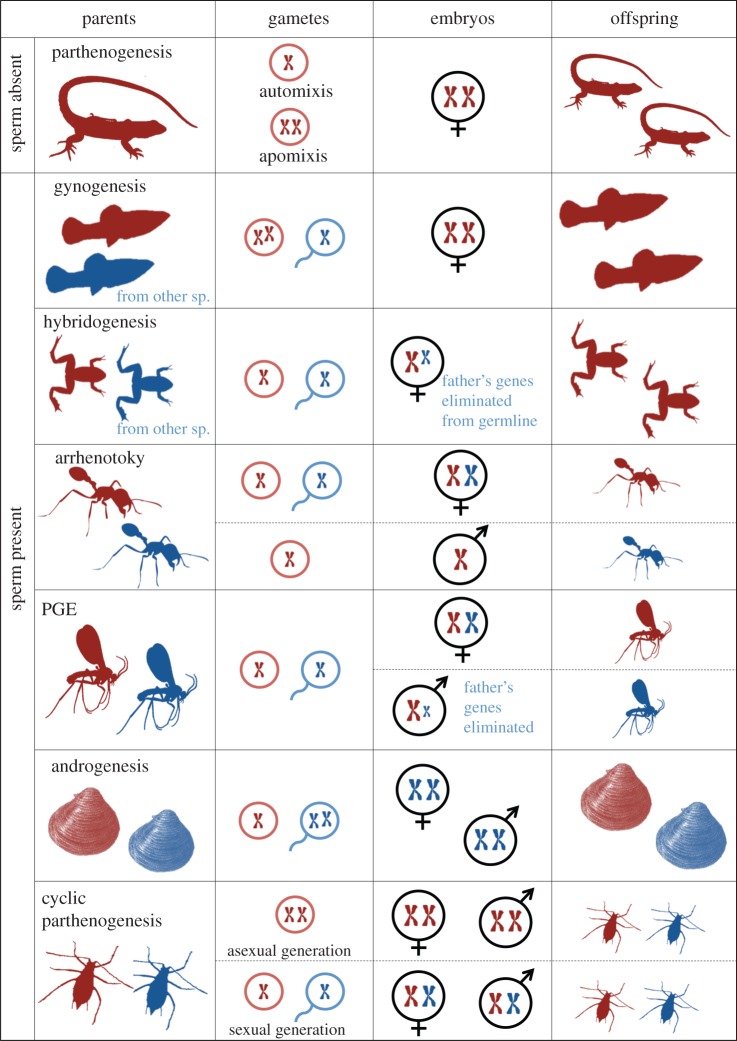
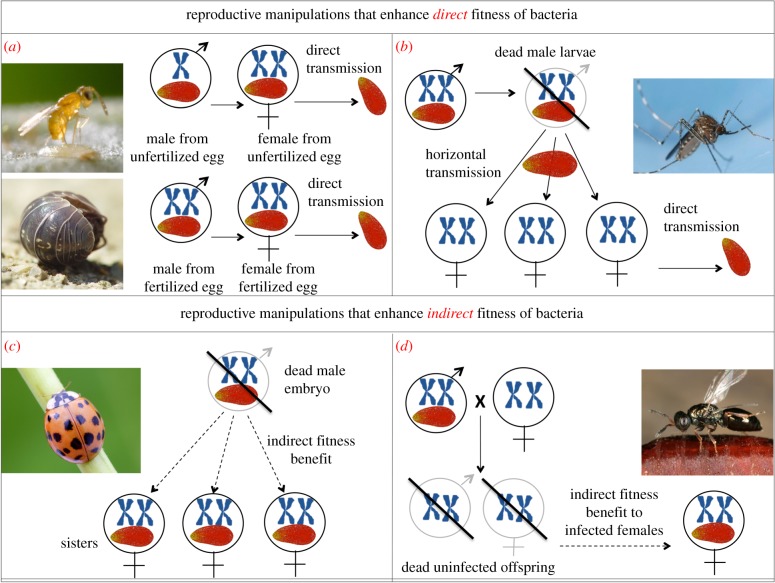
Genetic conflict may have played an important role in the evolution of novel genetic systems. The ancestral system of eumendelian genetics is highly symmetrical. Those derived from it (e.g. thelytokous parthenogenesis, haplodiploidy and parent-specific allele expression) are more asymmetrical in the genetic role played by maternal versus paternal alleles. These asymmetries may have arisen from maternal-paternal genetic conflict, or cytonuclear conflict, or from an interaction between them. Asymmetric genetic systems are much more common in terrestrial and freshwater taxa than in marine taxa. We suggest three reasons for this, based on the relative inhospitability of terrestrial environments to three types of organism: (i) pathogens-departure from the marine realm meant escape from many pathogens and parasites, reducing the need for sexual reproduction; (ii) symbionts-symbionts are no more important in the terrestrial realm than the marine realm but are more likely to be obligately intracellular and vertically transmitted, making them more likely to disrupt their host's genetic systems; (iii) Gametes and embryos-because neither gametes nor embryos can be shed into air as easily as into seawater, the mother's body is a more important environment for both types of organisms in the terrestrial realm than in the marine realm. This environment of asymmetric kinship (with neighbours more closely related by maternal alleles than by paternal alleles) may have helped to drive asymmetries in expression and transmission.
Keywords: Wolbachia; endosymbiont; genetic system; haplodiploidy; parthenogenesis.
Figures
Similar articles
-
Perspective: maternal kin groups and the origins of asymmetric genetic systems-genomic imprinting, haplodiploidy, and parthenogenesis.Evolution. 2006 Apr;60(4):631-42. Evolution. 2006. PMID: 16739447
-
Intragenomic conflict in populations infected by Parthenogenesis Inducing Wolbachia ends with irreversible loss of sexual reproduction.BMC Evol Biol. 2010 Jul 28;10:229. doi: 10.1186/1471-2148-10-229. BMC Evol Biol. 2010. PMID: 20667099 Free PMC article.
-
Can maternally transmitted endosymbionts facilitate the evolution of haplodiploidy?J Evol Biol. 2006 Jan;19(1):194-202. doi: 10.1111/j.1420-9101.2005.00974.x. J Evol Biol. 2006. PMID: 16405591
-
The evolution of alternative genetic systems in insects.Annu Rev Entomol. 2003;48:397-423. doi: 10.1146/annurev.ento.48.091801.112703. Epub 2002 Jun 4. Annu Rev Entomol. 2003. PMID: 12221039 Review.
-
The conflict theory of genomic imprinting: how much can be explained?Curr Top Dev Biol. 1998;40:255-93. doi: 10.1016/s0070-2153(08)60369-5. Curr Top Dev Biol. 1998. PMID: 9673853 Review.
Cited by
-
Males That Silence Their Father's Genes: Genomic Imprinting of a Complete Haploid Genome.Mol Biol Evol. 2021 May 19;38(6):2566-2581. doi: 10.1093/molbev/msab052. Mol Biol Evol. 2021. PMID: 33706381 Free PMC article.
-
The ghost sex-life of the paedogenetic beetle Micromalthus debilis.Sci Rep. 2016 Jun 7;6:27364. doi: 10.1038/srep27364. Sci Rep. 2016. PMID: 27270667 Free PMC article.
-
Conditional fitness benefits of the Rickettsia bacterial symbiont in an insect pest.Oecologia. 2016 Jan;180(1):169-79. doi: 10.1007/s00442-015-3436-x. Epub 2015 Sep 16. Oecologia. 2016. PMID: 26376661
-
Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflict.Genes (Basel). 2021 Jul 27;12(8):1136. doi: 10.3390/genes12081136. Genes (Basel). 2021. PMID: 34440310 Free PMC article. Review.
-
Gene-rich X chromosomes implicate intragenomic conflict in the evolution of bizarre genetic systems.Proc Natl Acad Sci U S A. 2022 Jun 7;119(23):e2122580119. doi: 10.1073/pnas.2122580119. Epub 2022 Jun 2. Proc Natl Acad Sci U S A. 2022. PMID: 35653559 Free PMC article.
References
-
- Leigh EG., Jr 1971. Adaptation and diversity. San Francisco, CA: Freeman, Cooper.
-
- Burt A, Trivers R. 2006. Genes in conflict: the biology of selfish genetic elements. Cambridge, MA: Harvard University Press.
-
- Bell G. 1982. The masterpiece of nature. Berkeley, CA: University of California Press.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
