

War and peace: social interactions in infections
- PMID: 24686936
- PMCID: PMC3982666
- DOI: 10.1098/rstb.2013.0365
War and peace: social interactions in infections
Abstract
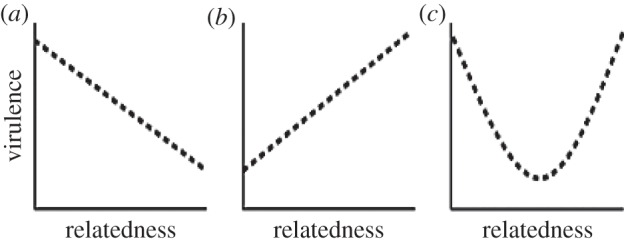
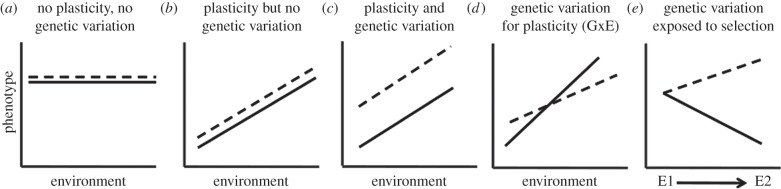
One of the most striking facts about parasites and microbial pathogens that has emerged in the fields of social evolution and disease ecology in the past few decades is that these simple organisms have complex social lives, indulging in a variety of cooperative, communicative and coordinated behaviours. These organisms have provided elegant experimental tests of the importance of relatedness, kin discrimination, cooperation and competition, in driving the evolution of social strategies. Here, we briefly review the social behaviours of parasites and microbial pathogens, including their contributions to virulence, and outline how inclusive fitness theory has helped to explain their evolution. We then take a mechanistically inspired 'bottom-up' approach, discussing how key aspects of the ways in which parasites and pathogens exploit hosts, namely public goods, mobile elements, phenotypic plasticity, spatial structure and multi-species interactions, contribute to the emergent properties of virulence and transmission. We argue that unravelling the complexities of within-host ecology is interesting in its own right, and also needs to be better incorporated into theoretical evolution studies if social behaviours are to be understood and used to control the spread and severity of infectious diseases.
Keywords: ecology; inclusive fitness; plasticity; relatedness; transmission; virulence.
Figures

References
-
- West SA, Diggle SP, Buckling A, Gardner A, Griffin AS. 2007. The social lives of microbes. Annu. Rev. Ecol. Evol. Syst. 38, 53–77. ( 10.1146/annurev.ecolsys.38.091206.095740) - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous