

Oxidative stress induces mitochondrial dysfunction in a subset of autistic lymphoblastoid cell lines
- PMID: 24690598
- PMCID: PMC4012280
- DOI: 10.1038/tp.2014.15
Oxidative stress induces mitochondrial dysfunction in a subset of autistic lymphoblastoid cell lines
Erratum in
-
Oxidative stress induces mitochondrial dysfunction in a subset of autistic lymphoblastoid cell lines.Transl Psychiatry. 2015 Mar 10;5(3):e526. doi: 10.1038/tp.2015.29. Transl Psychiatry. 2015. PMID: 25756809 Free PMC article. No abstract available.
Abstract
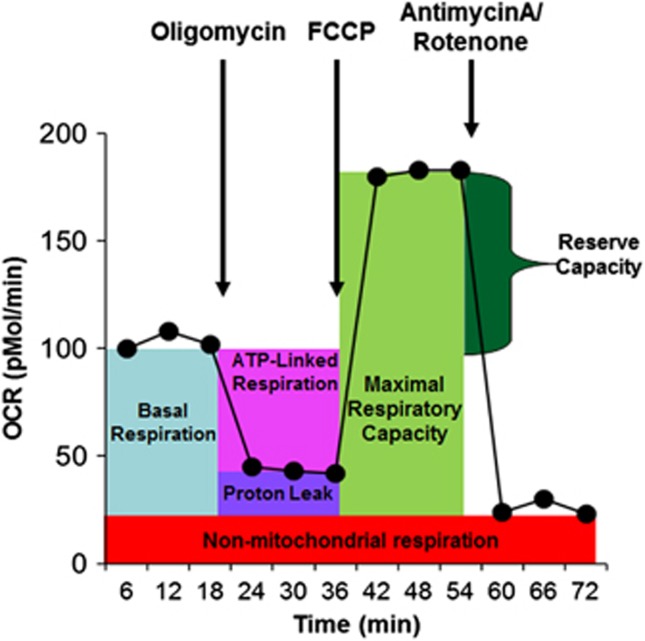
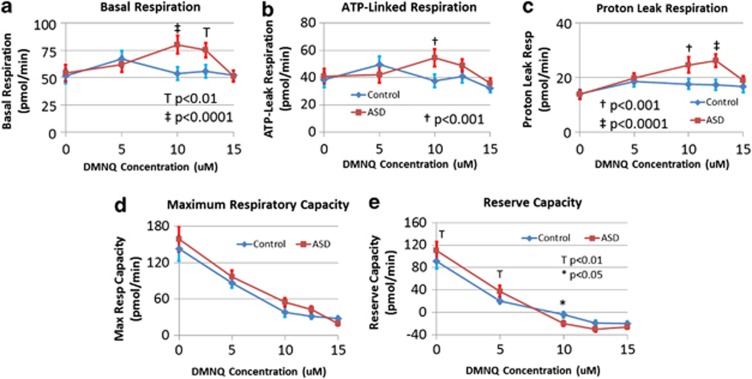
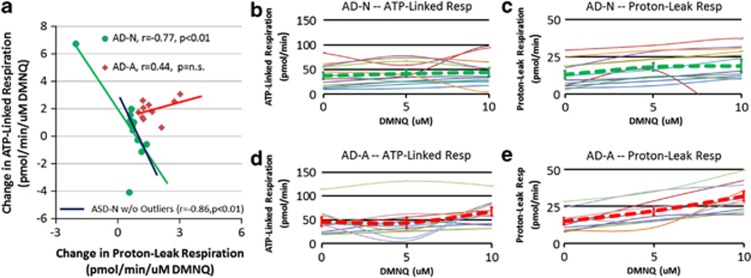
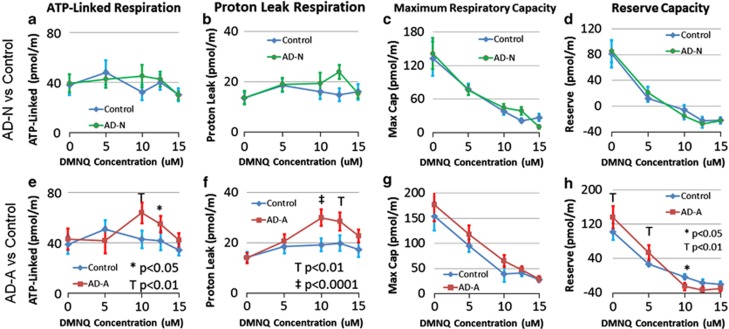
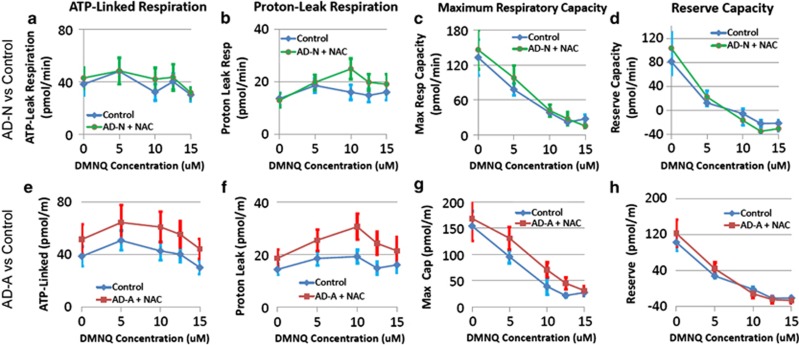
There is an increasing recognition that mitochondrial dysfunction is associated with autism spectrum disorders. However, little attention has been given to the etiology of mitochondrial dysfunction and how mitochondrial abnormalities might interact with other physiological disturbances such as oxidative stress. Reserve capacity is a measure of the ability of the mitochondria to respond to physiological stress. In this study, we demonstrate, for the first time, that lymphoblastoid cell lines (LCLs) derived from children with autistic disorder (AD) have an abnormal mitochondrial reserve capacity before and after exposure to reactive oxygen species (ROS). Ten (44%) of 22 AD LCLs exhibited abnormally high reserve capacity at baseline and a sharp depletion of reserve capacity when challenged with ROS. This depletion of reserve capacity was found to be directly related to an atypical simultaneous increase in both proton-leak respiration and adenosine triphosphate-linked respiration in response to increased ROS in this AD LCL subgroup. In this AD LCL subgroup, 48-hour pretreatment with N-acetylcysteine, a glutathione precursor, prevented these abnormalities and improved glutathione metabolism, suggesting a role for altered glutathione metabolism associated with this type of mitochondrial dysfunction. The results of this study suggest that a significant subgroup of AD children may have alterations in mitochondrial function, which could render them more vulnerable to a pro-oxidant microenvironment as well as intrinsic and extrinsic sources of ROS such as immune activation and pro-oxidant environmental toxins. These findings are consistent with the notion that AD is caused by a combination of genetic and environmental factors.
Figures
References
-
- American Psychiatric Association Diagnostic and Statistical Manual of Mental Disorders,4th edn. American Psychiatric Association: Washington, DC, USA; 1994
-
- Autism and Developmental Disabilities Monitoring Network Surveillance Year Principal Investigators, Centers for Disease Control and Prevention Prevalence of autism spectrum disorders—Autism and Developmental Disabilities Monitoring Network, United States, 2006. MMWR Surveill Summ. 2009;58:1–20. - PubMed
-
- Fowler BA, Woods JS. Ultrastructural and biochemical changes in renal mitochondria during chronic oral methyl mercury exposure: the relationship to renal function. Exp Mol Pathol. 1977;27:403–412. - PubMed
-
- Shenker BJ, Guo TL, O I, Shapiro IM. Induction of apoptosis in human T-cells by methyl mercury: temporal relationship between mitochondrial dysfunction and loss of reductive reserve. Toxicol Appl Pharmacol. 1999;157:23–35. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
