

Altered lipid homeostasis in Sertoli cells stressed by mild hyperthermia
- PMID: 24690895
- PMCID: PMC3972175
- DOI: 10.1371/journal.pone.0091127
Altered lipid homeostasis in Sertoli cells stressed by mild hyperthermia
Abstract
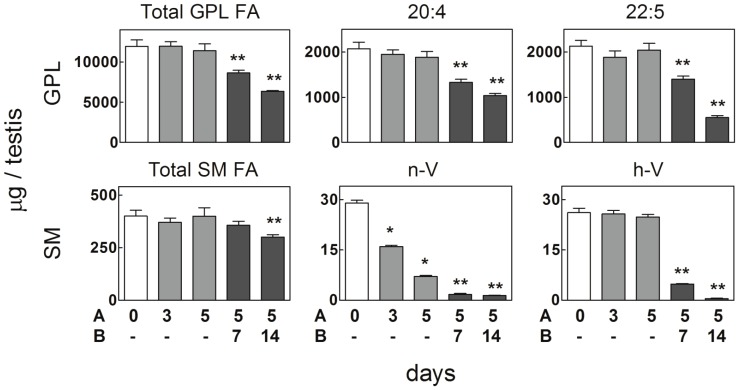
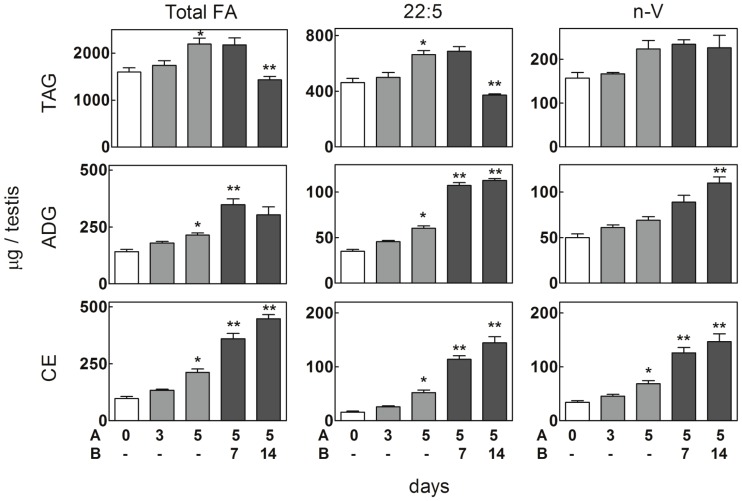
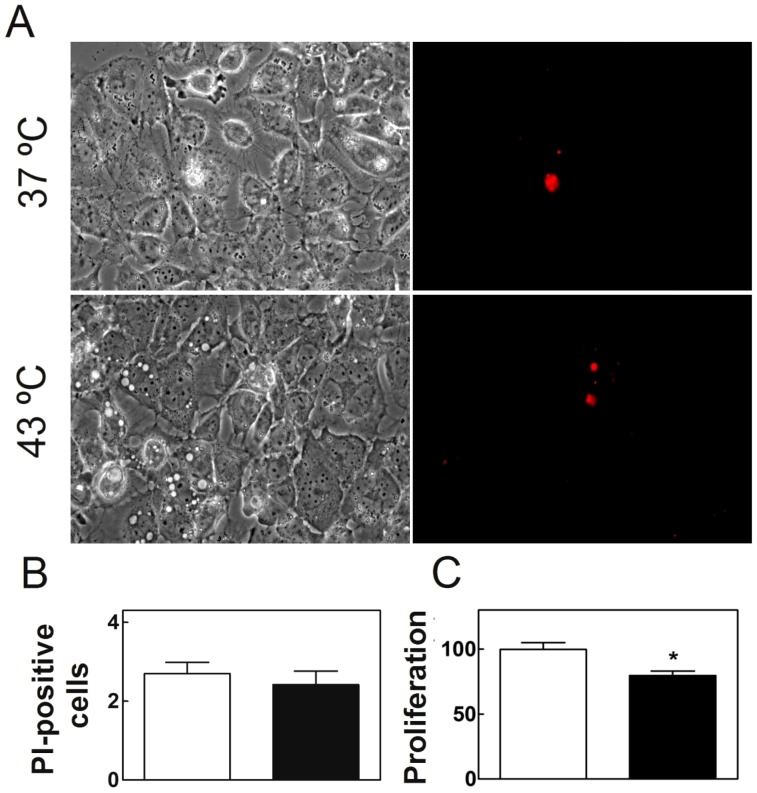
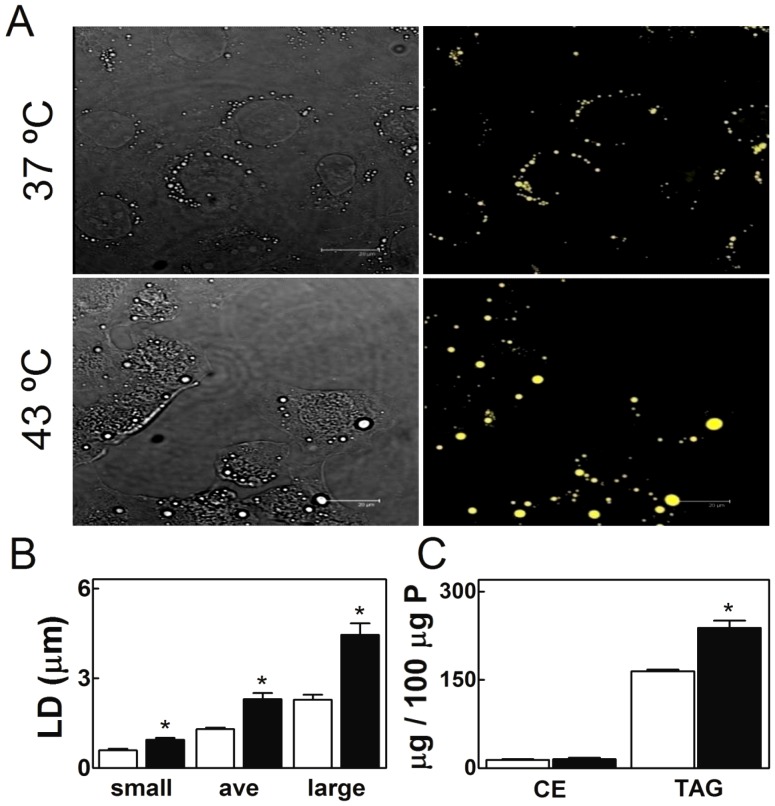
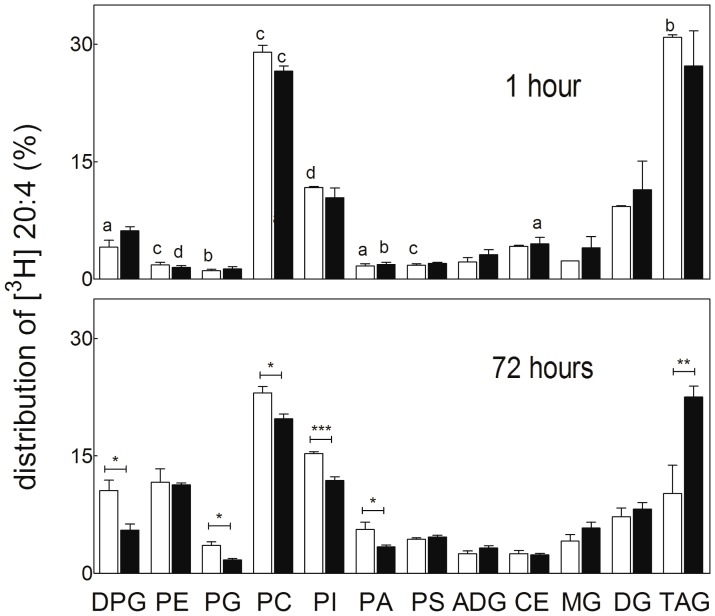
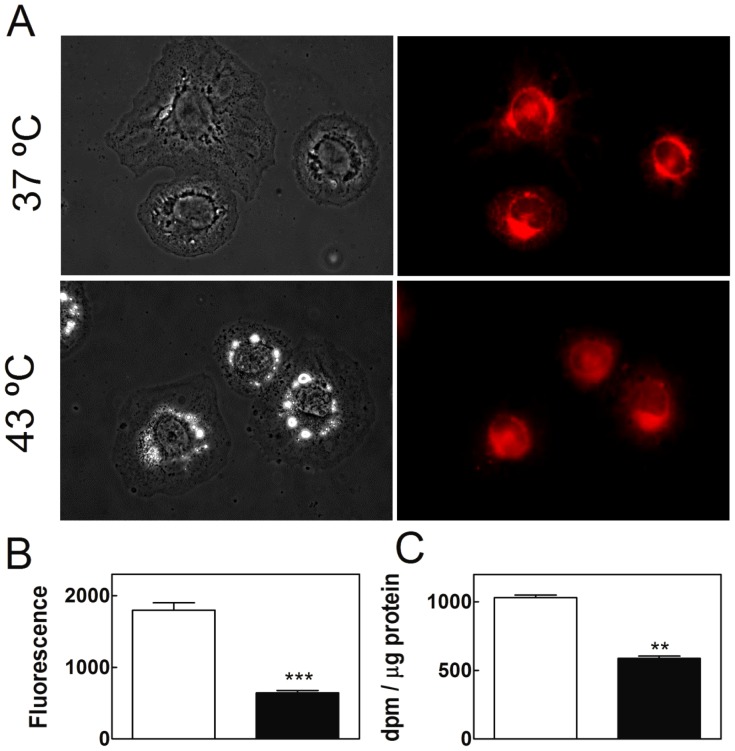
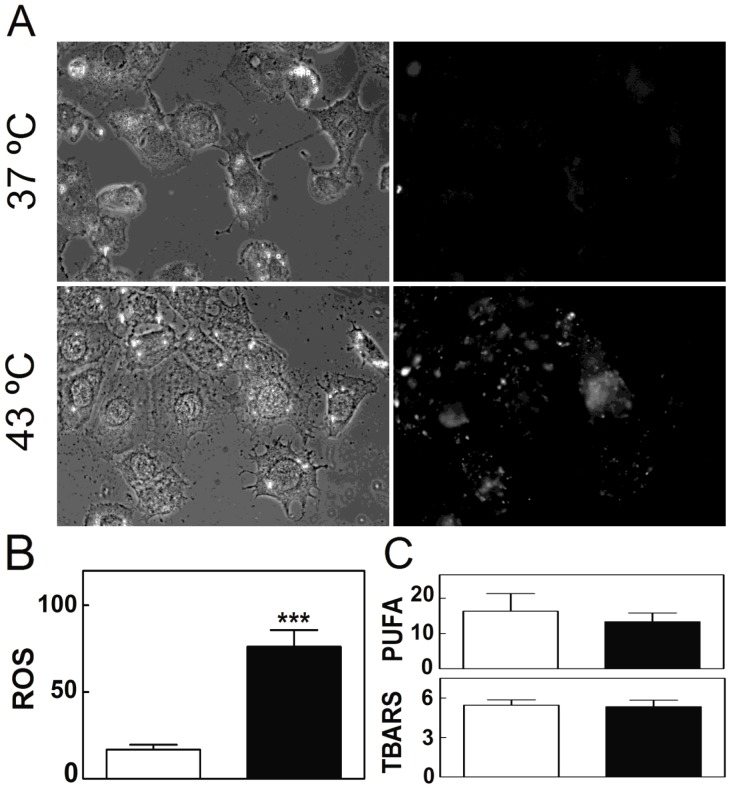
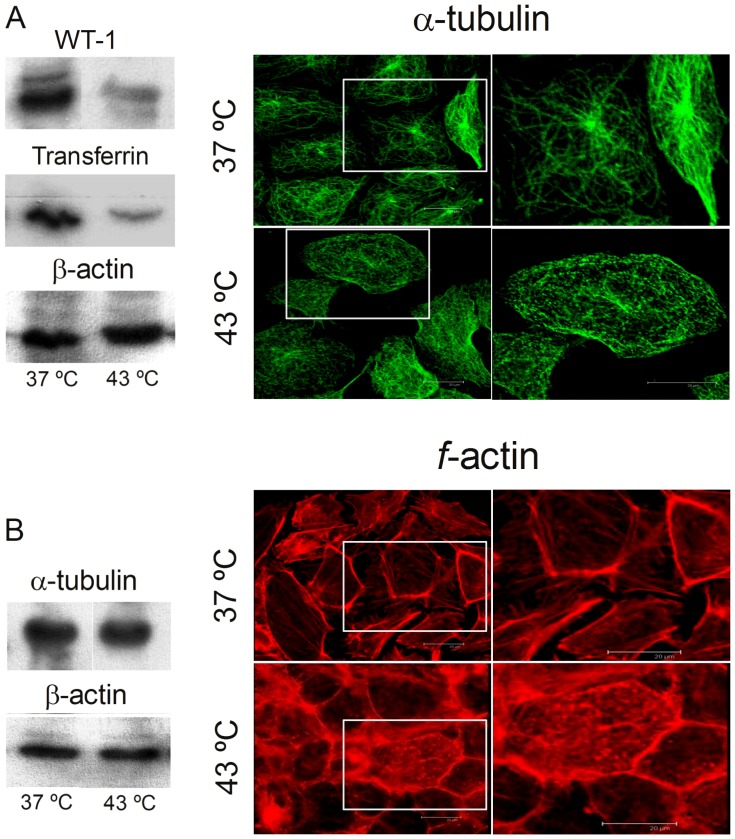
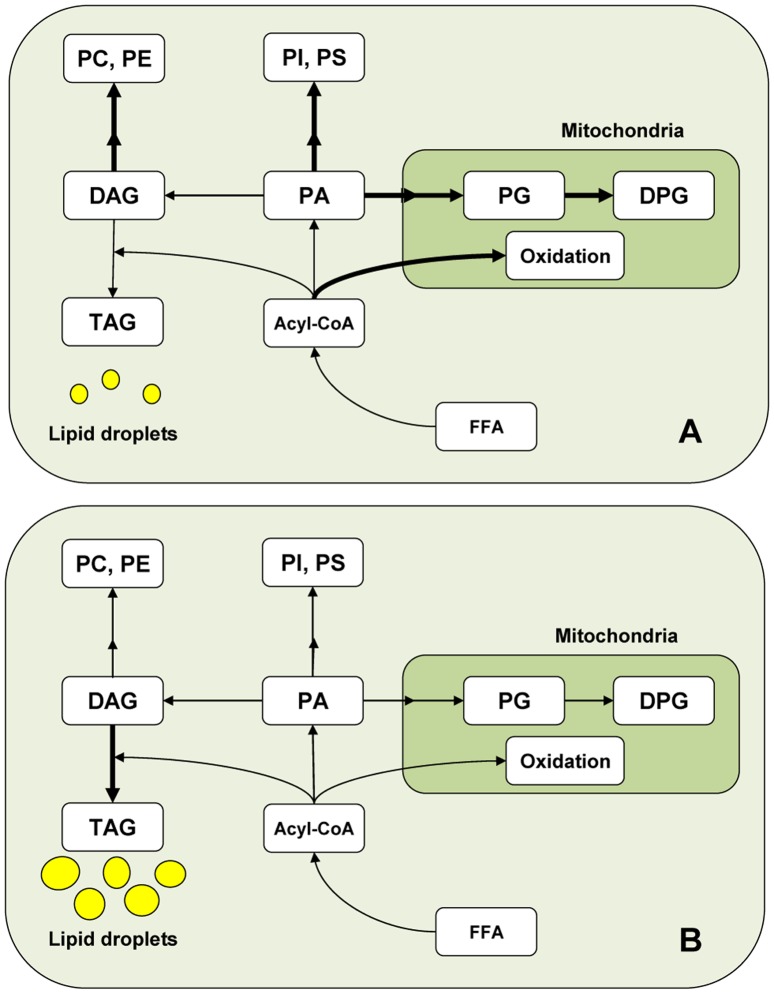
Spermatogenesis is known to be vulnerable to temperature. Exposures of rat testis to moderate hyperthermia result in loss of germ cells with survival of Sertoli cells (SC). Because SC provide structural and metabolic support to germ cells, our aim was to test the hypothesis that these exposures affect SC functions, thus contributing to germ cell damage. In vivo, regularly repeated exposures (one of 15 min per day, once a day during 5 days) of rat testes to 43 °C led to accumulation of neutral lipids. This SC-specific lipid function took 1-2 weeks after the last of these exposures to be maximal. In cultured SC, similar daily exposures for 15 min to 43 °C resulted in significant increase in triacylglycerol levels and accumulation of lipid droplets. After incubations with [3H]arachidonate, the labeling of cardiolipin decreased more than that of other lipid classes. Another specifically mitochondrial lipid metabolic function, fatty acid oxidation, also declined. These lipid changes suggested that temperature affects SC mitochondrial physiology, which was confirmed by significantly increased degrees of membrane depolarization and ROS production. This concurred with reduced expression of two SC-specific proteins, transferrin, and Wilms' Tumor 1 protein, markers of SC secretion and differentiation functions, respectively, and with an intense SC cytoskeletal perturbation, evident by loss of microtubule network (α-tubulin) and microfilament (f-actin) organization. Albeit temporary and potentially reversible, hyperthermia-induced SC structural and metabolic alterations may be long-lasting and/or extensive enough to respond for the decreased survival of the germ cells they normally foster.
Conflict of interest statement
Figures
References
-
- Moore CR (1924) The behavior of the germinal epithelium in testis grafts and in experimental cryptorchid testes (rat and guinea pig). Science 59: 41–44. - PubMed
-
- Setchell BP (1998) The Parkes Lecture. Heat and the testis. J Reprod Fertil 114: 179–194. - PubMed
-
- Yamamoto CM, Sinha Hikim AP, Huynh PN, Shapiro B, Lue Y, et al. (2000) Redistribution of Bax is an early step in an apoptotic pathway leading to germ cell death in rats, triggered by mild testicular hyperthermia. Biol Reprod 63: 1683–1690. - PubMed
-
- Rockett JC, Mapp FL, Garges JB, Luft JC, Mori C, et al. (2001) Effects of hyperthermia on spermatogenesis, apoptosis, gene expression, and fertility in adult male mice. Biol Reprod 65: 229–239. - PubMed
-
- Sinha Hikim AP, Swerdloff RS (1999) Hormonal and genetic control of germ cell apoptosis in the testis. Rev Reprod 4: 38–47. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
