

C-phycocyanin confers protection against oxalate-mediated oxidative stress and mitochondrial dysfunctions in MDCK cells
- PMID: 24691130
- PMCID: PMC3972226
- DOI: 10.1371/journal.pone.0093056
C-phycocyanin confers protection against oxalate-mediated oxidative stress and mitochondrial dysfunctions in MDCK cells
Erratum in
- PLoS One. 2014;9(7):e103361. Asokan, Devarajan [corrected to Devarajan, Asokan]
Abstract
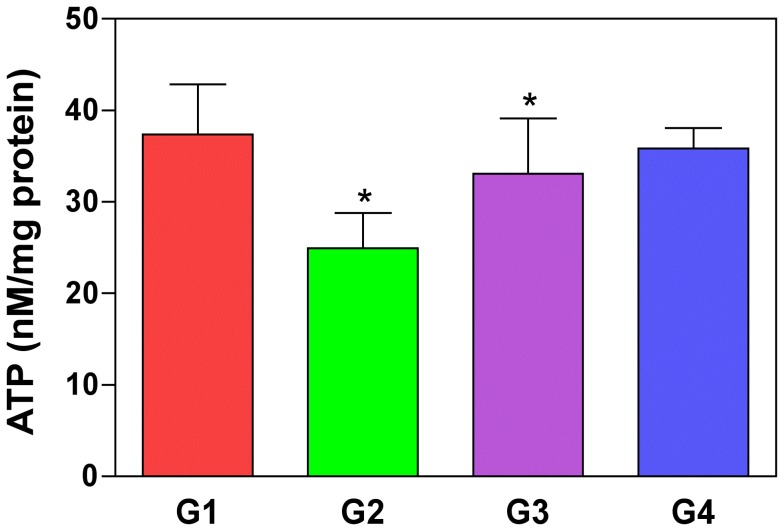
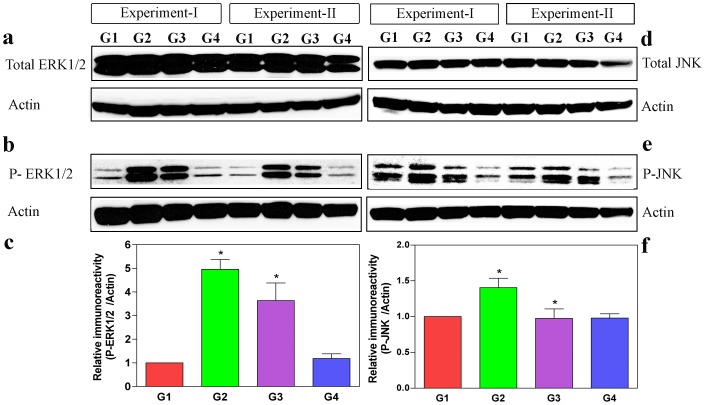
Oxalate toxicity is mediated through generation of reactive oxygen species (ROS) via a process that is partly dependent on mitochondrial dysfunction. Here, we investigated whether C-phycocyanin (CP) could protect against oxidative stress-mediated intracellular damage triggered by oxalate in MDCK cells. DCFDA, a fluorescence-based probe and hexanoyl-lysine adduct (HEL), an oxidative stress marker were used to investigate the effect of CP on oxalate-induced ROS production and membrane lipid peroxidation (LPO). The role of CP against oxalate-induced oxidative stress was studied by the evaluation of mitochondrial membrane potential by JC1 fluorescein staining, quantification of ATP synthesis and stress-induced MAP kinases (JNK/SAPK and ERK1/2). Our results revealed that oxalate-induced cells show markedly increased ROS levels and HEL protein expression that were significantly decreased following pre-treatment with CP. Further, JC1 staining showed that CP pre-treatment conferred significant protection from mitochondrial membrane permeability and increased ATP production in CP-treated cells than oxalate-alone-treated cells. In addition, CP treated cells significantly decreased the expression of phosphorylated JNK/SAPK and ERK1/2 as compared to oxalate-alone-treated cells. We concluded that CP could be used as a potential free radical-scavenging therapeutic strategy against oxidative stress-associated diseases including urolithiasis.
Conflict of interest statement
Figures
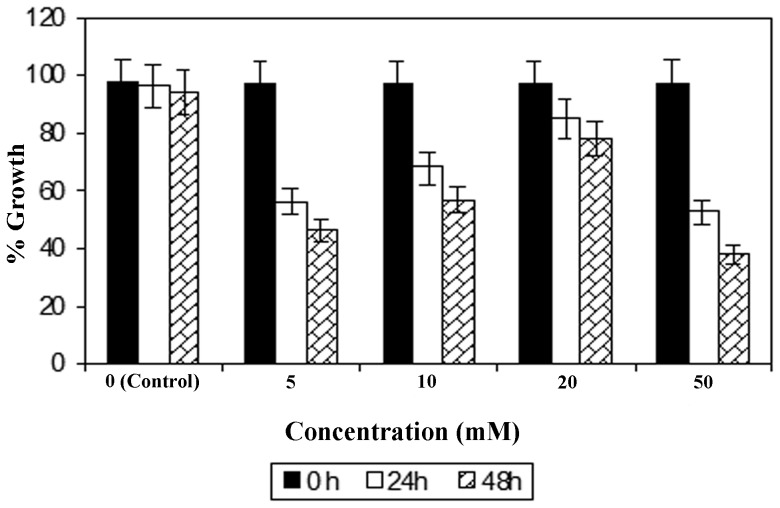
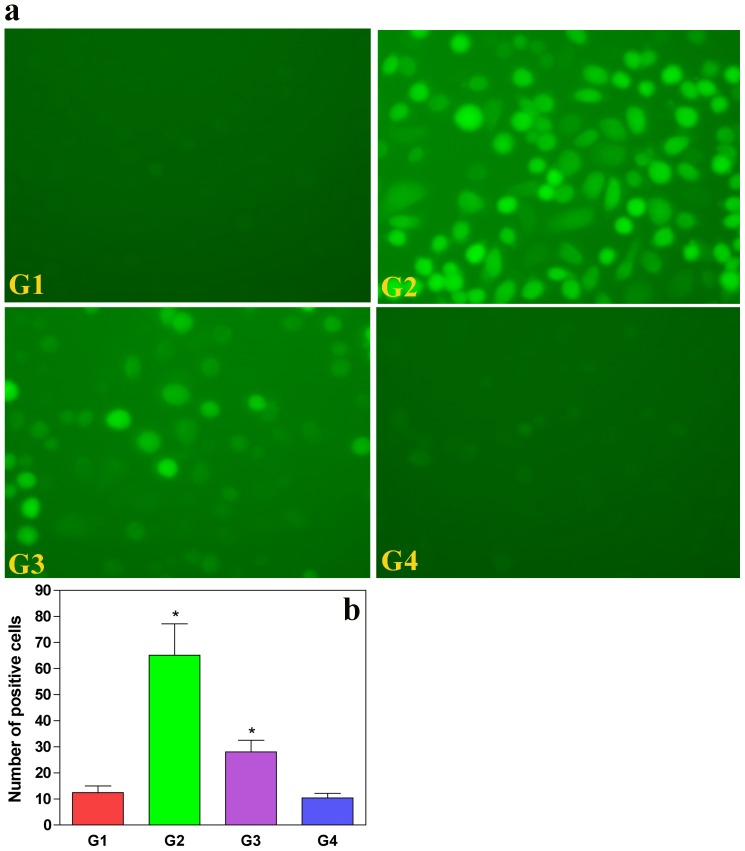
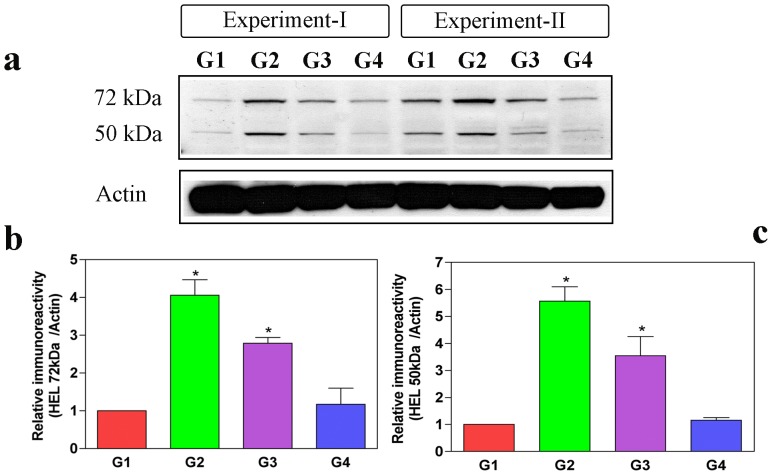
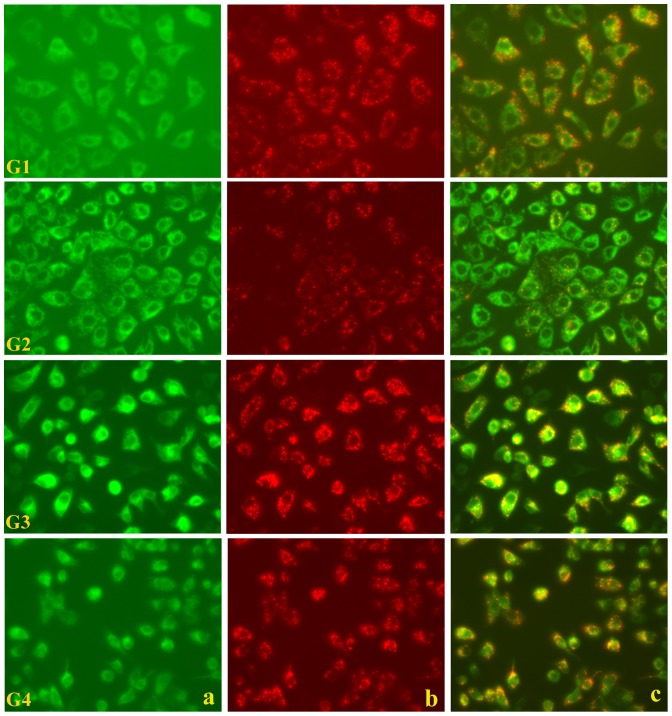
References
-
- Brzica H, Breljak D, Burckhardt BC, Burckhardt G, Sabolic I (2013) Oxalate: from the environment to kidney stones. Arh Hig Rada Toksikol 64: 609–630. - PubMed
-
- Johri N, Cooper B, Robertson W, Choong S, Rickards D, et al. (2010) An update and practical guide to renal stone management. Nephron Clin Pract 116: c159–171. - PubMed
-
- Thamilselvan V, Menon M, Thamilselvan S (2014) Oxalate at physiological urine concentrations induces oxidative injury in renal epithelial cells: Effect of α-tocopherol and ascorbic acid. BJU Int doi: 10.1111/bju.12642 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
