

Transporters MCT8 and OATP1C1 maintain murine brain thyroid hormone homeostasis
- PMID: 24691440
- PMCID: PMC4001533
- DOI: 10.1172/JCI70324
Transporters MCT8 and OATP1C1 maintain murine brain thyroid hormone homeostasis
Abstract
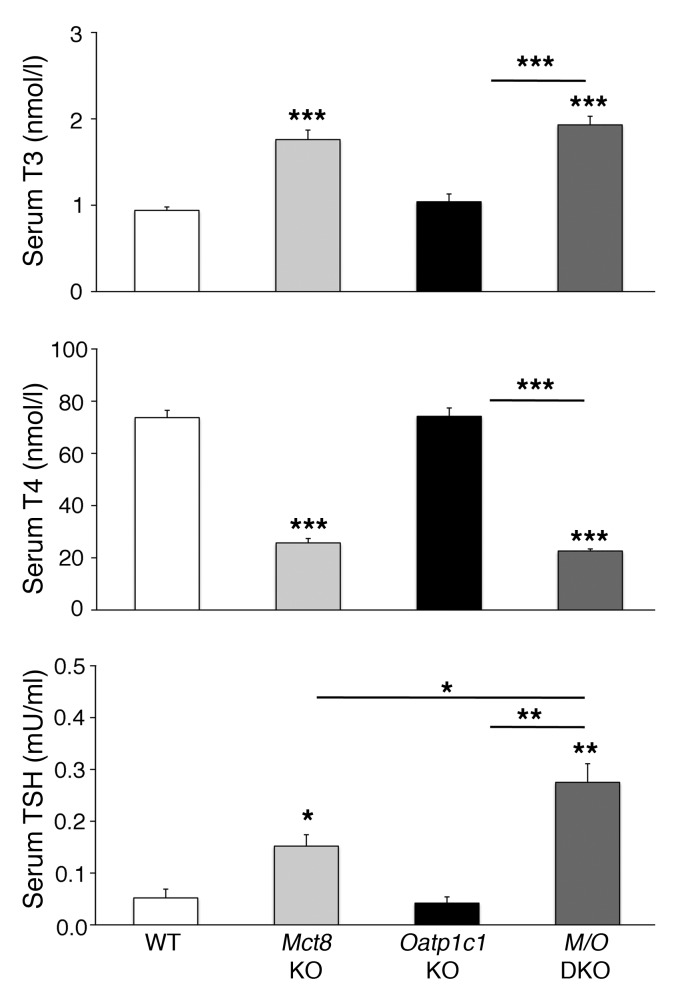
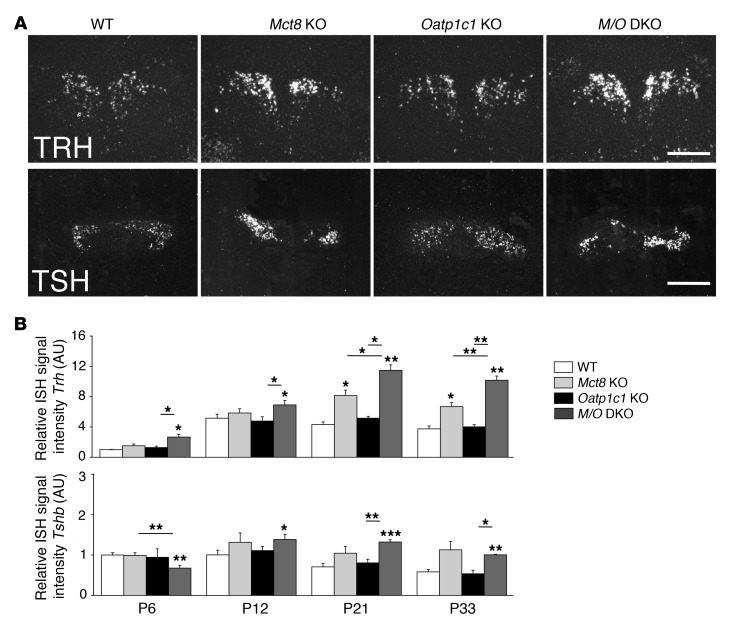
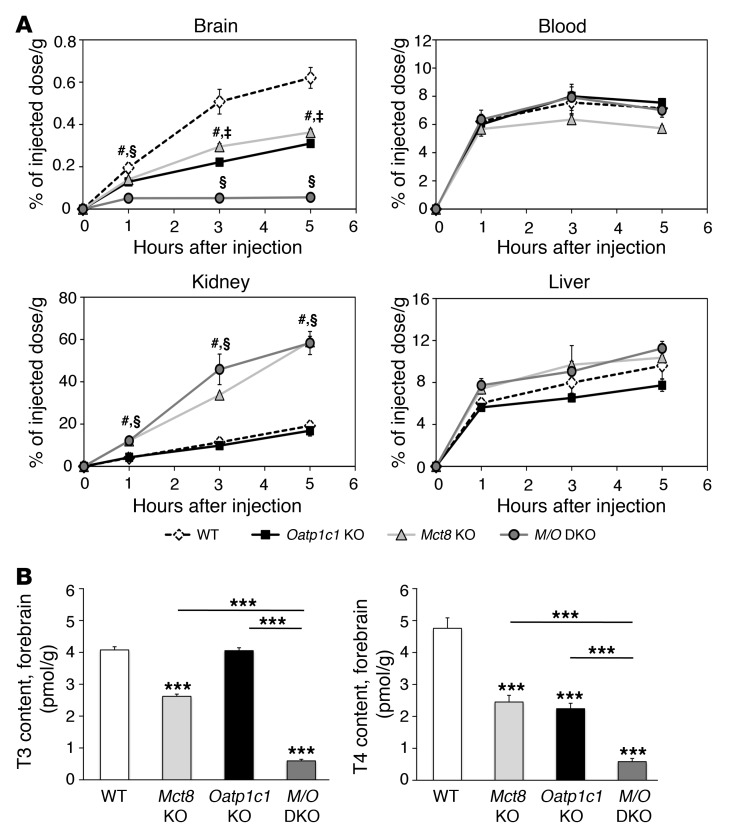
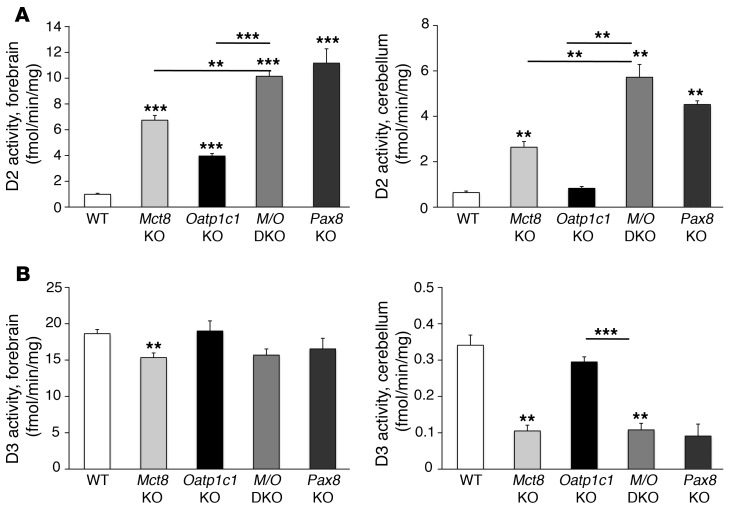
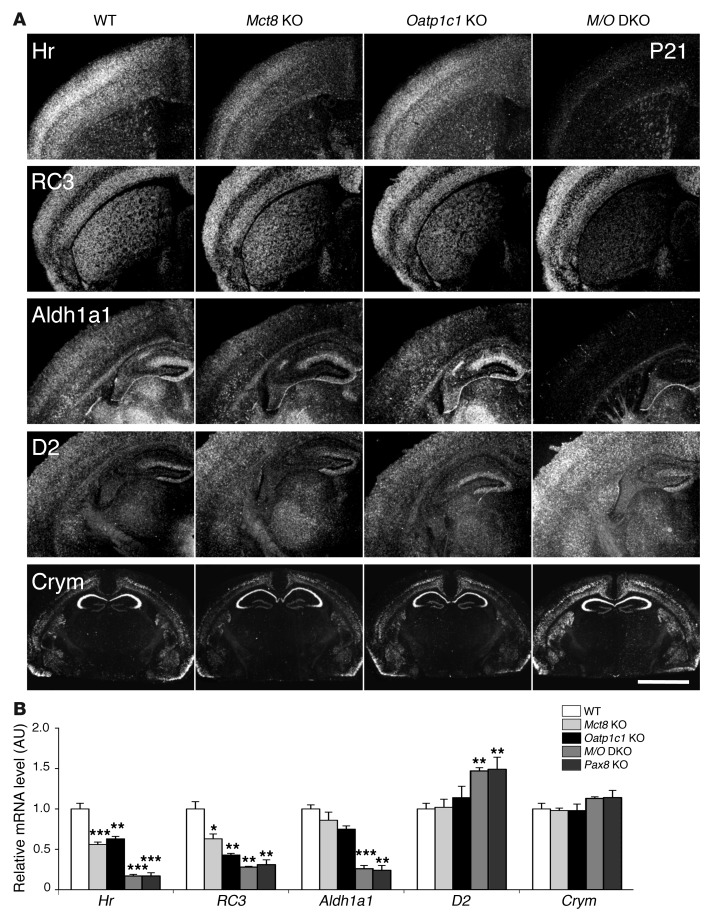
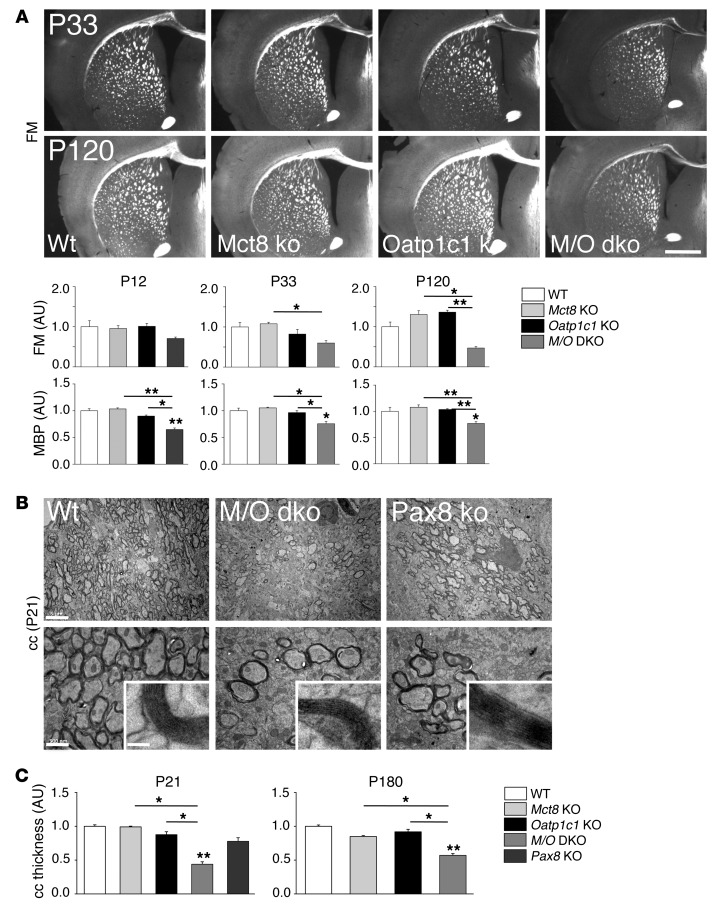
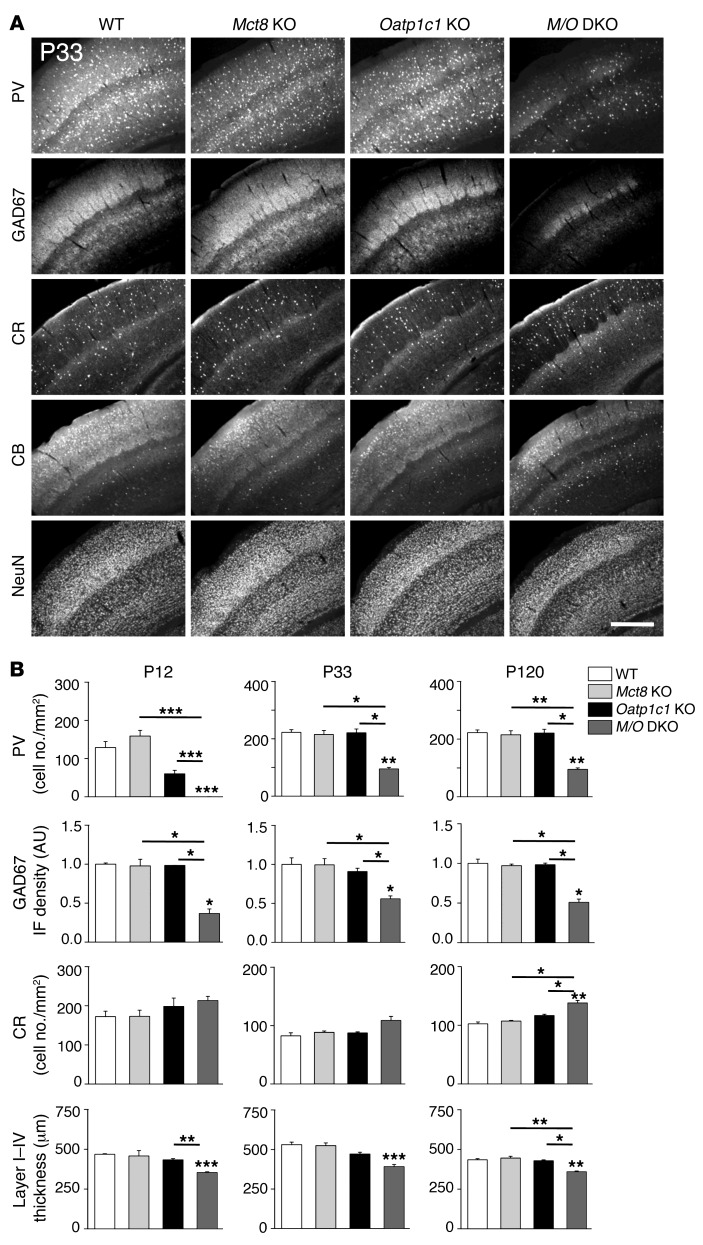
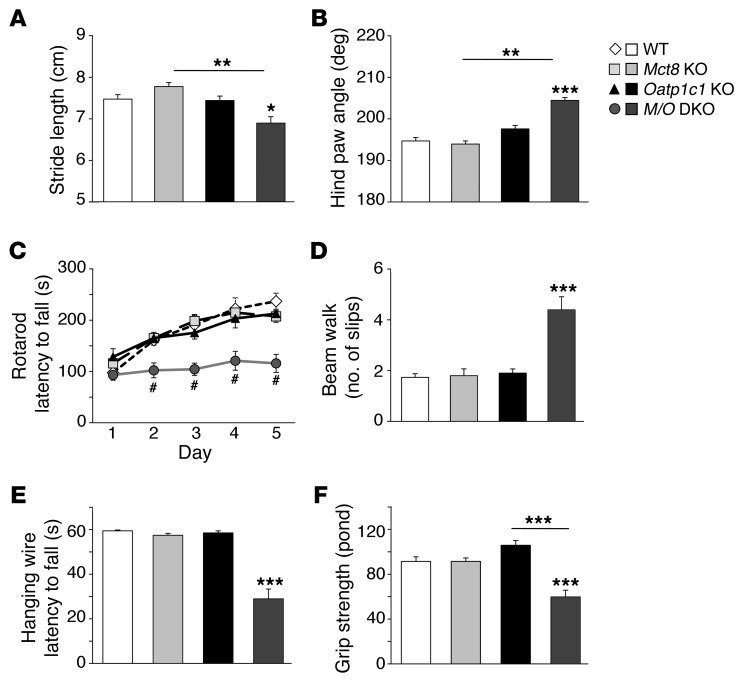
Allan-Herndon-Dudley syndrome (AHDS), a severe form of psychomotor retardation with abnormal thyroid hormone (TH) parameters, is linked to mutations in the TH-specific monocarboxylate transporter MCT8. In mice, deletion of Mct8 (Mct8 KO) faithfully replicates AHDS-associated endocrine abnormalities; however, unlike patients, these animals do not exhibit neurological impairments. While transport of the active form of TH (T3) across the blood-brain barrier is strongly diminished in Mct8 KO animals, prohormone (T4) can still enter the brain, possibly due to the presence of T4-selective organic anion transporting polypeptide (OATP1C1). Here, we characterized mice deficient for both TH transporters, MCT8 and OATP1C1 (Mct8/Oatp1c1 DKO). Mct8/Oatp1c1 DKO mice exhibited alterations in peripheral TH homeostasis that were similar to those in Mct8 KO mice; however, uptake of both T3 and T4 into the brains of Mct8/Oatp1c1 DKO mice was strongly reduced. Evidence of TH deprivation in the CNS of Mct8/Oatp1c1 DKO mice included highly decreased brain TH content as well as altered deiodinase activities and TH target gene expression. Consistent with delayed cerebellar development and reduced myelination, Mct8/Oatp1c1 DKO mice displayed pronounced locomotor abnormalities. Intriguingly, differentiation of GABAergic interneurons in the cerebral cortex was highly compromised. Our findings underscore the importance of TH transporters for proper brain development and provide a basis to study the pathogenic mechanisms underlying AHDS.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
