

Review
doi: 10.1101/cshperspect.a018713.
Erasers of histone acetylation: the histone deacetylase enzymes
Affiliations
- PMID: 24691964
- PMCID: PMC3970420
- DOI: 10.1101/cshperspect.a018713
Item in Clipboard
Review
Erasers of histone acetylation: the histone deacetylase enzymes
Cold Spring Harb Perspect Biol.
.
Abstract
Histone deacetylases (HDACs) are enzymes that catalyze the removal of acetyl functional groups from the lysine residues of both histone and nonhistone proteins. In humans, there are 18 HDAC enzymes that use either zinc- or NAD(+)-dependent mechanisms to deacetylate acetyl lysine substrates. Although removal of histone acetyl epigenetic modification by HDACs regulates chromatin structure and transcription, deacetylation of nonhistones controls diverse cellular processes. HDAC inhibitors are already known potential anticancer agents and show promise for the treatment of many diseases.
Figures
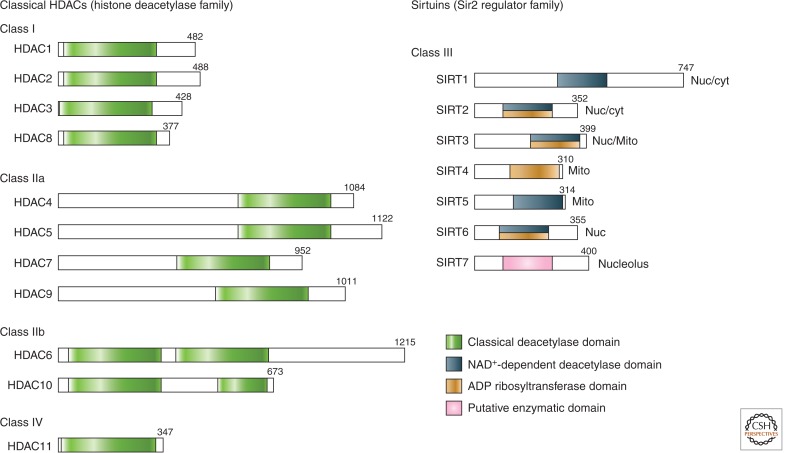
Domain organization of human HDACs. The total number of amino acid residues in each HDAC is shown on the right of each protein. Many HDACs have multiple isoforms and, for simplicity, only the longest isoform is shown. Enzymatic domains (or putative enzymatic domains) are shown in colors. Sirtuin localizations: Nuc, nuclear; cyt, cytoplasmic; Mito, mitochrondial.
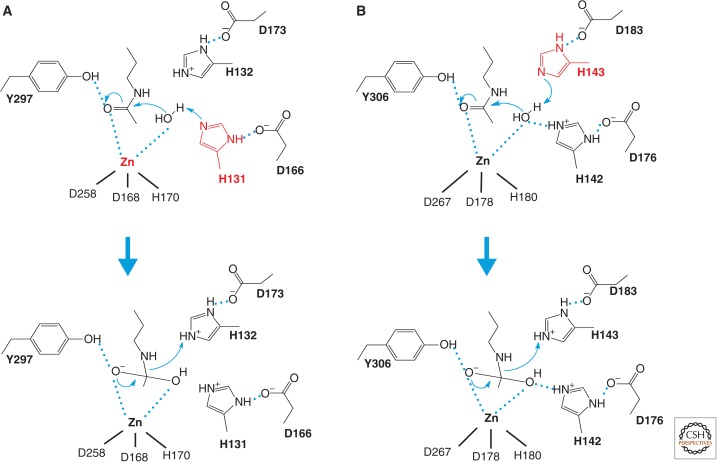
Catalytic mechanism of HDACs. Two models for the catalytic mechanism of the Zn-dependent HDAC reaction have been proposed. (A) A model proposed from the HDLP structure. The HDLP catalytic core consists of a tubular pocket, a zinc-binding site, and active-site residues (in bold) of a tyrosine (Y297) and two histidines (H131 and H132) that make hydrogen bonds to two aspartic acids (D166 and D173). One of these catalytic histidines (red) facilitates nucleophilic attack at the substrate carbonyl by activating a water molecule coordinated with the zinc ion. Initially, two tandem histidine residues (H131 and H132) were proposed to function as Asp-His charge relay systems, typical of serine proteases such as chymotrypsin and chymotrypsinogen in the enzyme reaction. The active site zinc ion is coordinated by three residues (two aspartic acids and one histidine). (B) A model proposed from the HDAC8 structure, in which the other histidine residue (red) plays an essential role in the electron transfer. Hydrogen bond interactions are drawn in dotted lines.
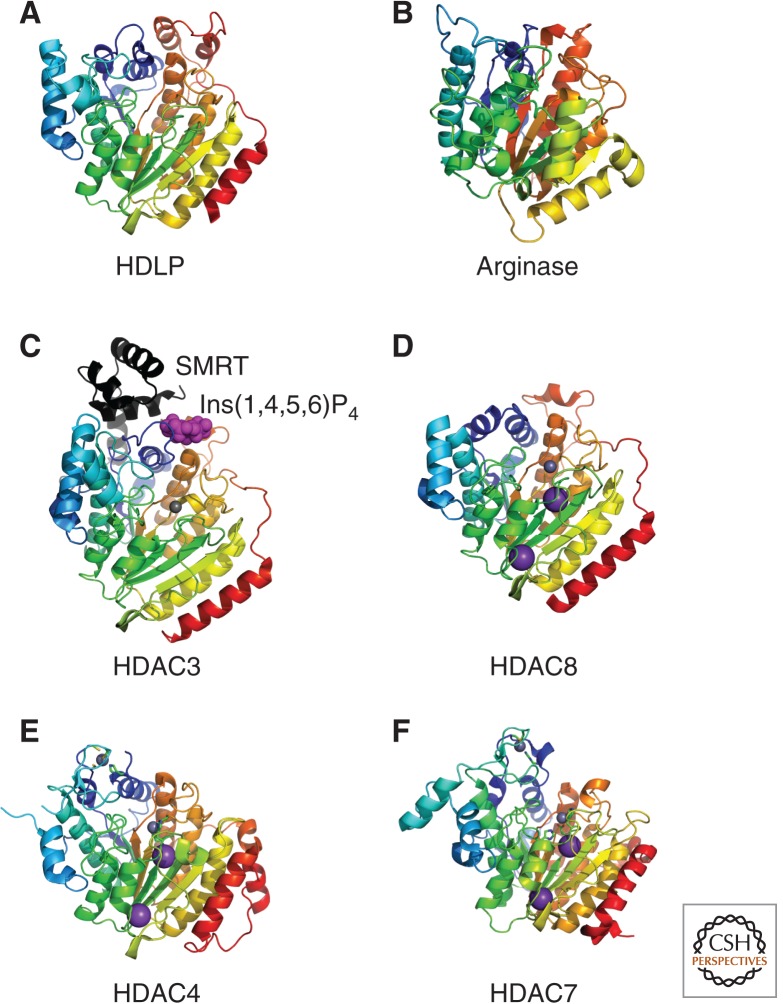
Structure of Class I and II HDACs. Crystal structures of the arginase/deacetylase superfamily of proteins are illustrated. Metal ions are represented as space filled spheres: red, gray, and violet spheres indicate Mn, Zn, and K atoms, respectively. (A) Aquifex aeolicus HDLP (1C3P), (B) rat arginase (PDB ID: 1RLA), (C) human Class I HDAC3 (4A69) in complex with Ins(1,4,5,6)P4 (highlighted by magenta space-filled spheres) and the deacetylase activation domain from the human SMRT corepressor (depicted with a ribbon model in black), (D) human Class I HDAC8 (3F07), (E) human Class IIa HDAC4 (2VQW), and (F) human Class IIa HDAC7 (3C0Y).
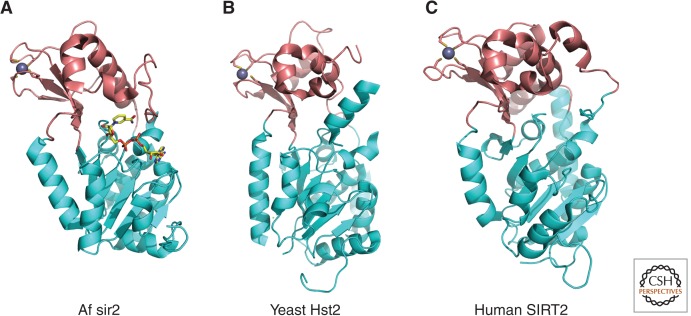
Overall structure of sirtuins (Class III). Crystal structures of the sirtuin family of proteins are illustrated as cartoons highlighting the large Rossmann-fold domains (cyan) and small zinc-binding domains (brown). (A) Archaeoglobus fulgidus sir2 (PDB ID: 1ICI). NAD+ is drawn in stick model form in which yellow, blue, red, and orange represent C, N, O, and P atoms, respectively. (B) Yeast Hst2 (1Q14). (C) Human SIRT2 (1J8F). Zinc ions are represented as space-filled spheres in gray.
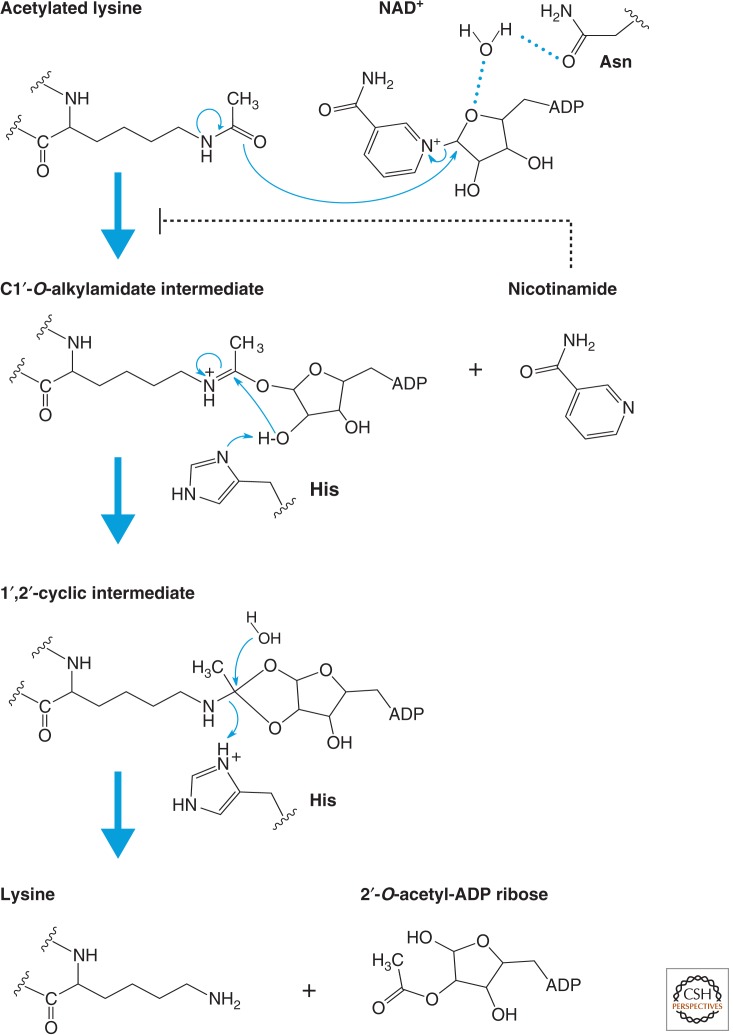
Catalytic mechanism of sirtuins (Class III). Proposed mechanism of the NAD+-dependent deacetylase reaction. The first step of the reaction involves nucleophilic addition of the acetamide oxygen to the C1′ position of the nicotinamide ribose to form a C1′-O-alkylamidate intermediate and free nicotinamide. Next, the 2′-hydroxy group of the NAD+ ribose is activated by an active site histidine residue that, in turn, attacks the C1′-O-alkylamidate to form the 1′,2′-cyclic intermediate. The 1′,2′-cyclic intermediate is then attacked by an activated water molecule resulting in the formation of deacetylated lysine and 2′-O-acetyl-ADP ribose. 2′-O-acetyl-ADP ribose can be readily converted to 3′-O-acetyl-ADP ribose in aqueous solution by nonenzymatic intramolecular transesterification. Thus, nicotinamide, the deacetylated peptide, and a mixture of 2′- and 3′-O-acetyl-ADP ribose are the final reaction products.
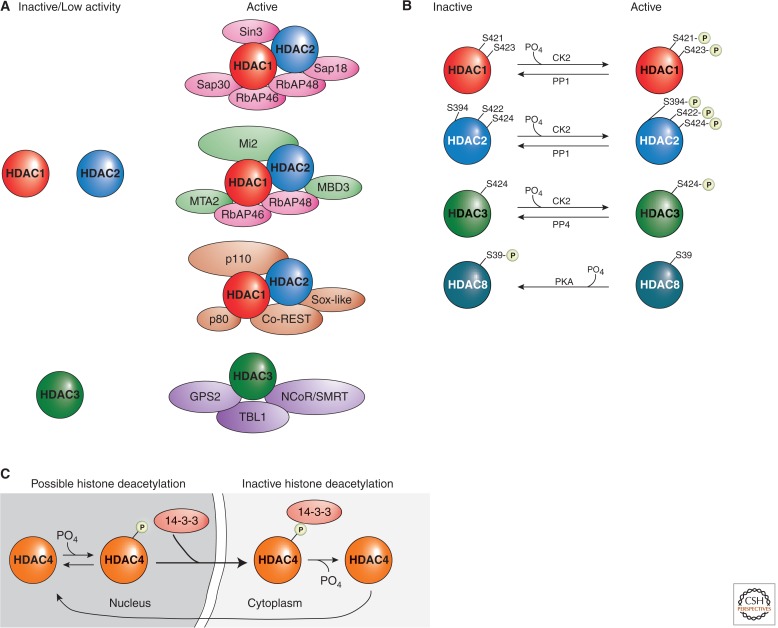
Examples of HDAC regulation. (A) Class I HDACs are commonly regulated by protein complex formations. HDAC1, HDAC2, and HDAC3 possess low enzymatic activities when in isolation, and their activities increase significantly when present in holoenzyme complexes. (B) The activities of Class I HDACs are modulated by phosphorylation and dephosphorylation. In general, phosphorylation activates HDAC1, HDAC2, and HDAC3, although it represses HDAC8 activities. (C) Phosphorylation of a Class II HDAC, HDAC4, promotes its interaction with the 14-3-3 protein and, subsequently, changes its localization. Multiple residues (S246, S467, and S632 on HDAC4, and corresponding conserved sites on HDAC5, HDAC7, and HDAC9) confer the HDAC–14-3-3 interactions.
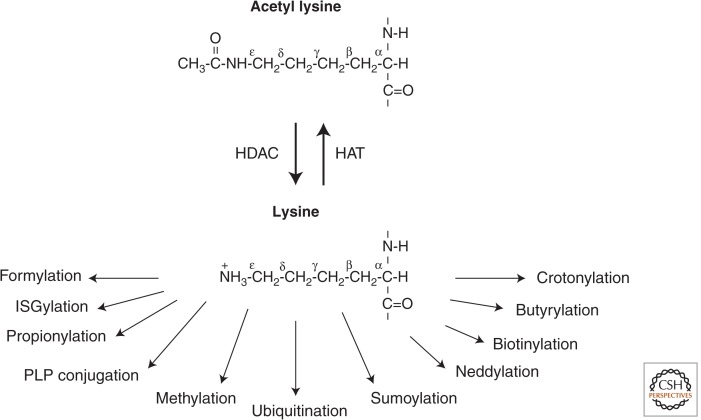
Examples of the many different potential posttranslational modifications on an ε-amino lysine subsequent to HDAC deacetylation.
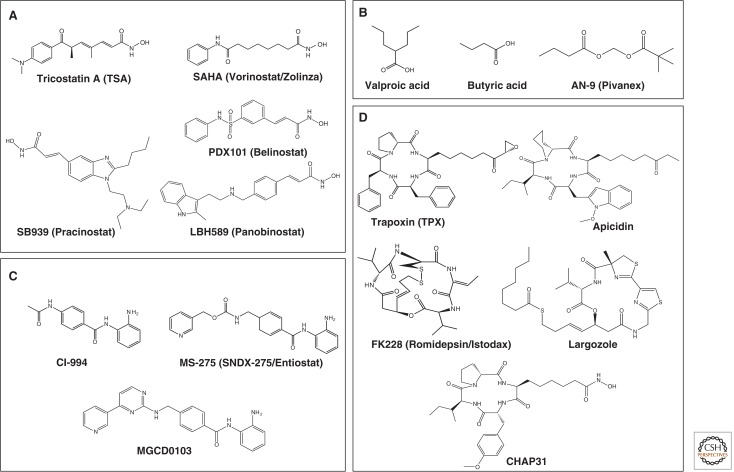
HDAC inhibitors. Four classes of HDAC inhibitors categorized according to their chemical structures are shown. (A) Hydroxamic acid, (B) short-chain fatty acid, (C) benzamide, and (D) cyclic peptide.
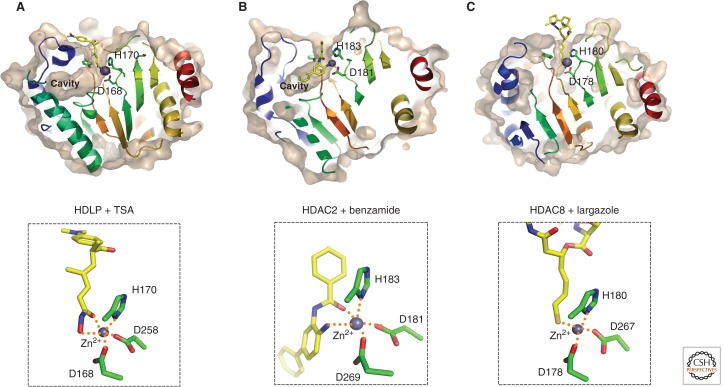
Crystal structure of HDAC proteins complexed with inhibitors. Molecular surfaces of the inhibitor-bound HDLP/HDACs cut at the level of the cavity are illustrated. (A) HDLP with TSA (PDB ID: 1C3R), (B) HDAC2 with benzamide (3MAX), and (C) HDAC8 with largazole (3RQD). The outer surfaces are depicted in light brown, and zinc atoms are shown in space-filled spheres in gray. Close-up views of the active sites are shown in the lower panels. Inhibitors are depicted in a stick model, in which yellow, blue, red, and gold represent C, N, O, and S atoms, respectively. Three Zn-coordinating residues (two aspartic acid and one histidine residue) are shown in a stick model. Hydrogen bond interactions are drawn in dotted lines.
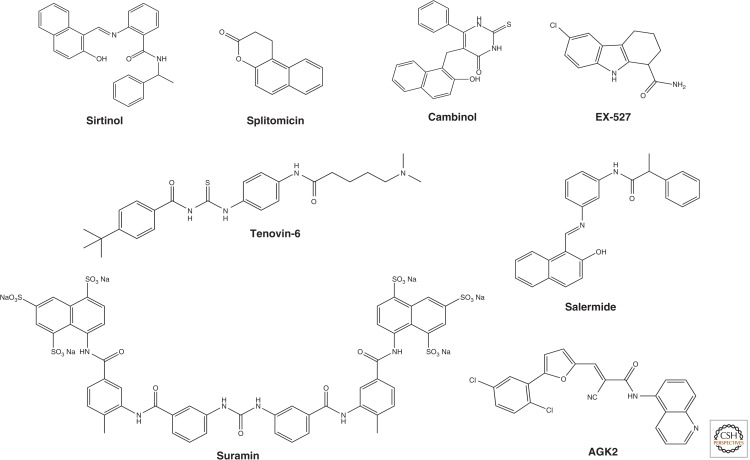
Sirtuin inhibitors. Sirtinol and splitomicin are identified as the first small molecule sirtuin inhibitors that affect telomere silencing in yeast. Cambinol is a splitomicin-related β-naphthol that is more stable than splitomicin and increases p53 acetylation showing antitumor activity in BCL6-expressing Burkitt’s lymphoma. Salermide was designed based on the structure of sirtinol by molecular modeling and inhibits both SIRT1 and SIRT2 more effectively than sirtinol. Tenovin-1 and its water-soluble analog tenovin-6 induce p53 acetylation; their cellular targets were determined to be SIRT1 and SIRT2. A high-throughput screen revealed a number of indole compounds including EX-527, which selectively inhibits SIRT1 over SIRT2. Kinetic analysis suggests that EX-527 binds to the nicotinamide-binding site. AGK2, which was reported as a SIRT2 selective inhibitor, shows more than 10-fold selective inhibition relative to SIRT1 and SIRT3. Suramin, which was originally developed for treating trypanosomiasis and onchocerciasis, inhibits the NAD+-dependent deacetylase activity of sirtuins by inducing sirtuin dimerization.
References
-
- Allis D, Jenuwein T, Reinberg D 2014. Overview and concepts. Cold Spring Harb Perspect Biol 10.1101/cshperspect.a018739 - DOI
-
- Asaba T, Suzuki T, Ueda R, Tsumoto H, Nakagawa H, Miyata N 2009. Inhibition of human sirtuins by in situ generation of an acetylated lysine-ADP-ribose conjugate. J Am Chem Soc 131: 6989–6996 - PubMed
-
- Avalos JL, Boeke JD, Wolberger C 2004. Structural basis for the mechanism and regulation of Sir2 enzymes. Mol Cell 13: 639–648 - PubMed
-
- Ayer DE 1999. Histone deacetylases: Transcriptional repression with SINers and NuRDs. Trends Cell Biol 9: 193–198 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources