

RIG-I detects mRNA of intracellular Salmonella enterica serovar Typhimurium during bacterial infection
- PMID: 24692634
- PMCID: PMC3977358
- DOI: 10.1128/mBio.01006-14
RIG-I detects mRNA of intracellular Salmonella enterica serovar Typhimurium during bacterial infection
Abstract
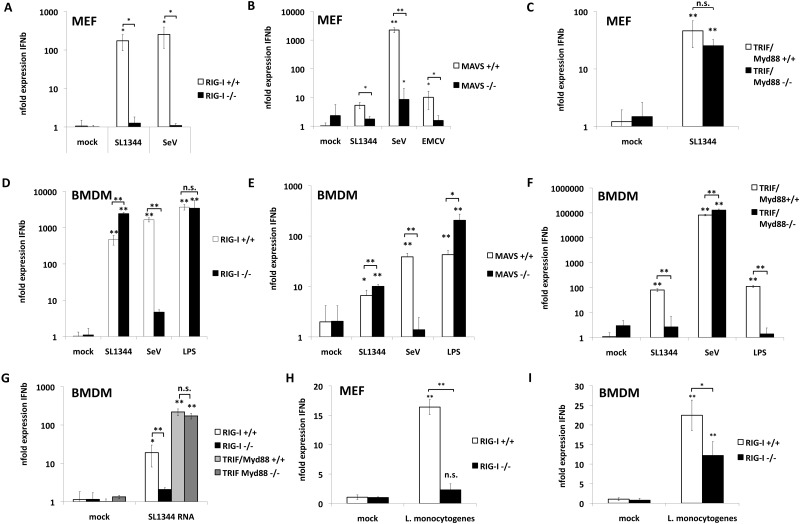
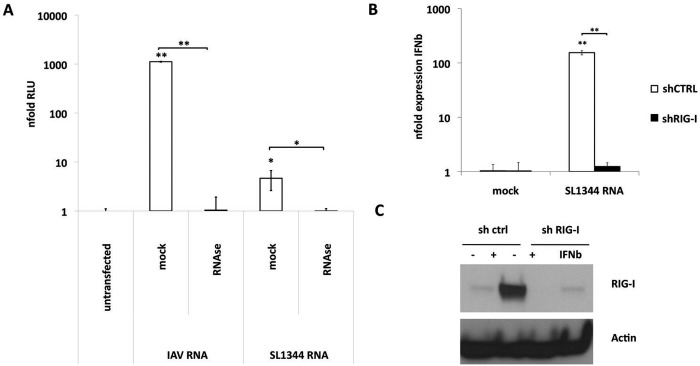
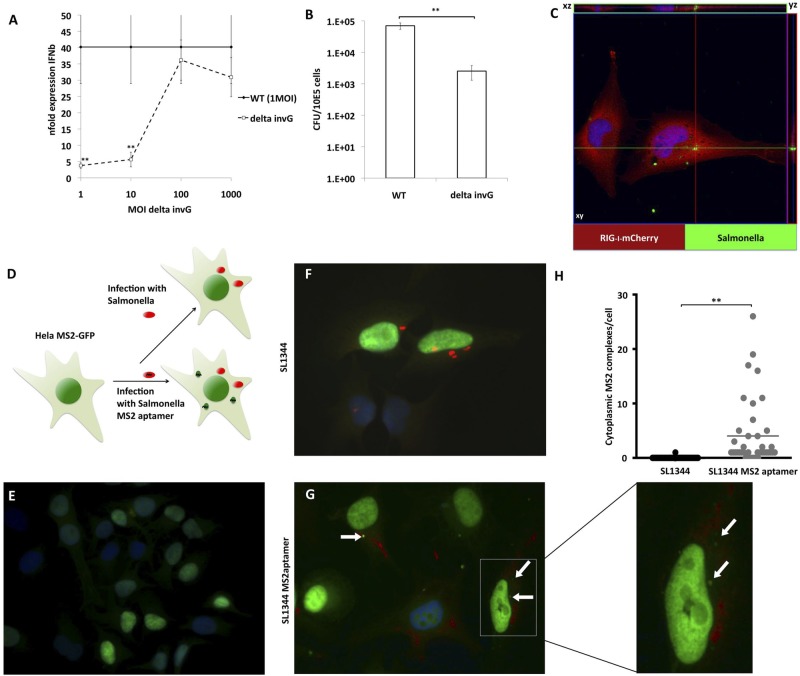
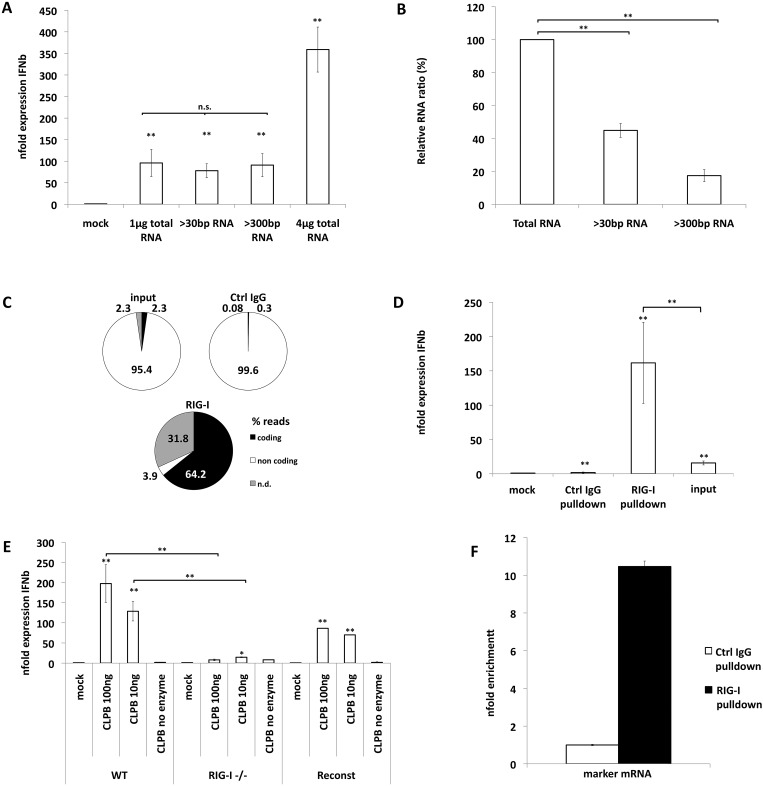
The cytoplasmic helicase RIG-I is an established sensor for viral 5'-triphosphorylated RNA species. Recently, RIG-I was also implicated in the detection of intracellular bacteria. However, little is known about the host cell specificity of this process and the bacterial pathogen-associated molecular pattern (PAMP) that activates RIG-I. Here we show that RNA of Salmonella enterica serovar Typhimurium activates production of beta interferon in a RIG-I-dependent fashion only in nonphagocytic cells. In phagocytic cells, RIG-I is obsolete for detection of Salmonella infection. We further demonstrate that Salmonella mRNA reaches the cytoplasm during infection and is thus accessible for RIG-I. The results from next-generation sequencing analysis of RIG-I-associated RNA suggest that coding bacterial mRNAs represent the activating PAMP. IMPORTANCE S. Typhimurium is a major food-borne pathogen. After fecal-oral transmission, it can infect epithelial cells in the gut as well as immune cells (mainly macrophages, dendritic cells, and M cells). The innate host immune system relies on a growing number of sensors that detect pathogen-associated molecular patterns (PAMPs) to launch a first broad-spectrum response to invading pathogens. Successful detection of a given pathogen depends on colocalization of host sensors and PAMPs as well as potential countermeasures of the pathogen during infection. RIG-I-like helicases were mainly associated with detection of RNA viruses. Our work shows that S. Typhimurium is detected by RIG-I during infection specifically in nonimmune cells.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
