

Minireview: Extrapituitary prolactin: an update on the distribution, regulation, and functions
- PMID: 24694306
- PMCID: PMC5414853
- DOI: 10.1210/me.2013-1349
Minireview: Extrapituitary prolactin: an update on the distribution, regulation, and functions
Abstract
Prolactin (PRL) is an important hormone with many diverse functions. Although it is predominantly produced by lactrotrophs of the pituitary there are a number of other organs, cells, and tissues in which PRL is expressed and secreted. The impact of this extrapituitary PRL (ePRL) on localized metabolism and cellular functions is gaining widespread attention. In 1996, a comprehensive review on ePRL was published. However, since this time, there have been a number of advancements in ePRL research. This includes a greater understanding of the components of the control elements located within the superdistal promoter of the ePRL gene. Furthermore, several new sites of ePRL have been discovered, each under unique control by a range of transcription factors and elements. The functional role of ePRL at each of the expression sites also varies widely leading to gender and site bias. This review aims to provide an update to the research conducted on ePRL since the 1996 review. The focus is on new data concerning the sites of ePRL expression, its regulation, and its function within the organs in which it is expressed.
Figures
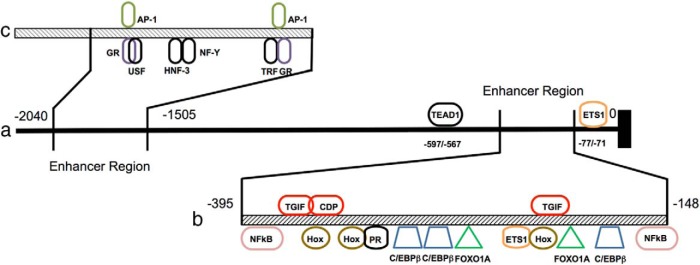
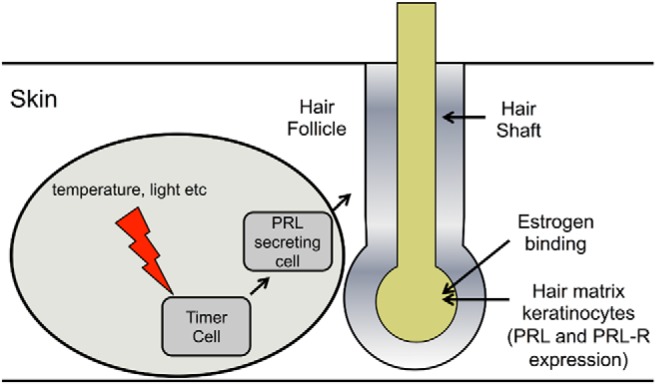
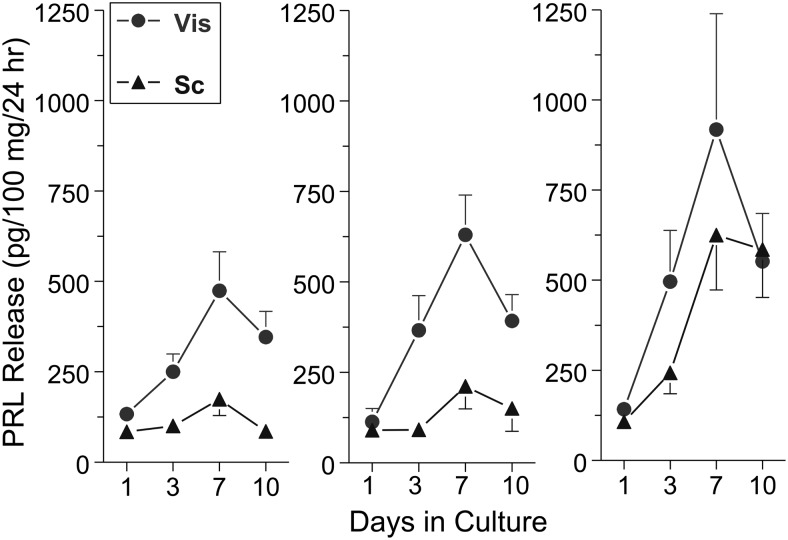
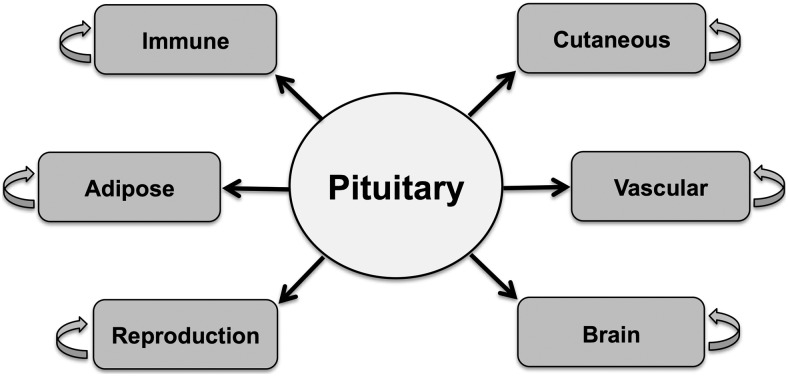
References
-
- Hiraoka Y, Tatsumi K, Shiozawa M, et al. A placenta-specific 5′ non-coding exon of human prolactin. Mol Cell Endocrinol. 1991;75:71–80. - PubMed
-
- Gellersen B, Kempf R, Telgmann R, DiMattia GE. Nonpituitary human prolactin gene transcription is independent of Pit-1 and differentially controlled in lymphocytes and in endometrial stroma. Mol Endocrinol. 1994;8:356–373. - PubMed
-
- Ben-Jonathan N, Mershon JL, Allen DL, Steinmetz RW. Extrapituitary prolactin: distribution, regulation, functions, and clinical aspects. Endocr Rev. 1996;17:639–669. - PubMed
-
- Arendt LM, Schuler LA. Transgenic models to study actions of prolactin in mammary neoplasia. J Mammary Gland Biol Neoplasia. 2008;13:29–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
