

Olfactory coding in the insect brain: data and conjectures
- PMID: 24698302
- PMCID: PMC4237541
- DOI: 10.1111/ejn.12558
Olfactory coding in the insect brain: data and conjectures
Abstract
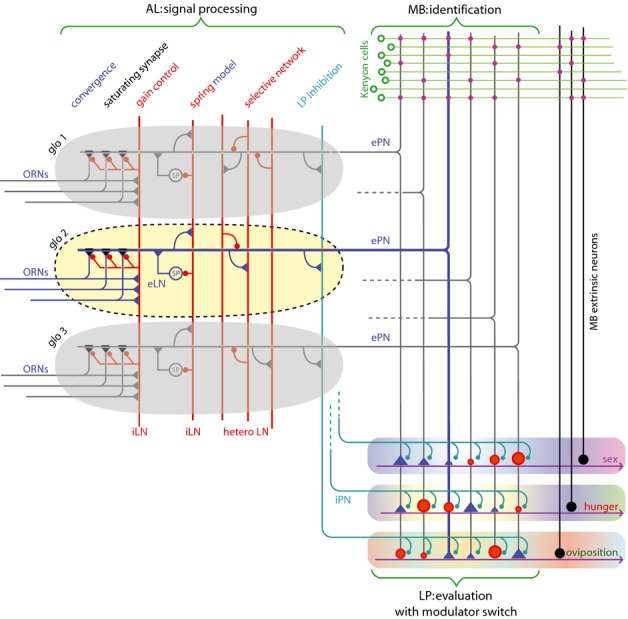
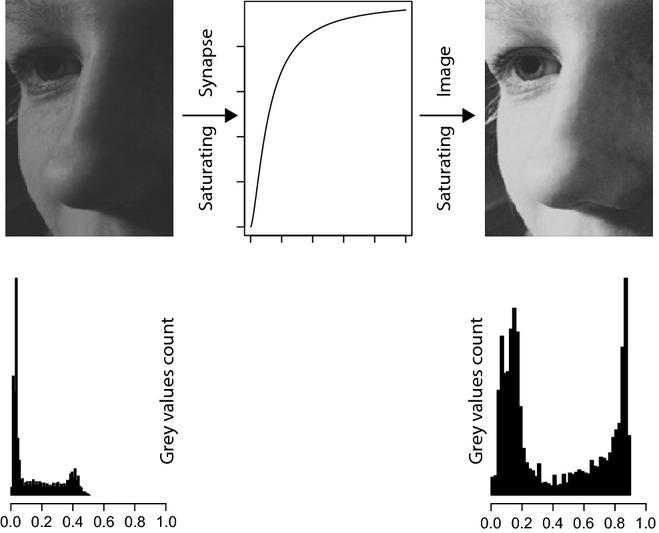
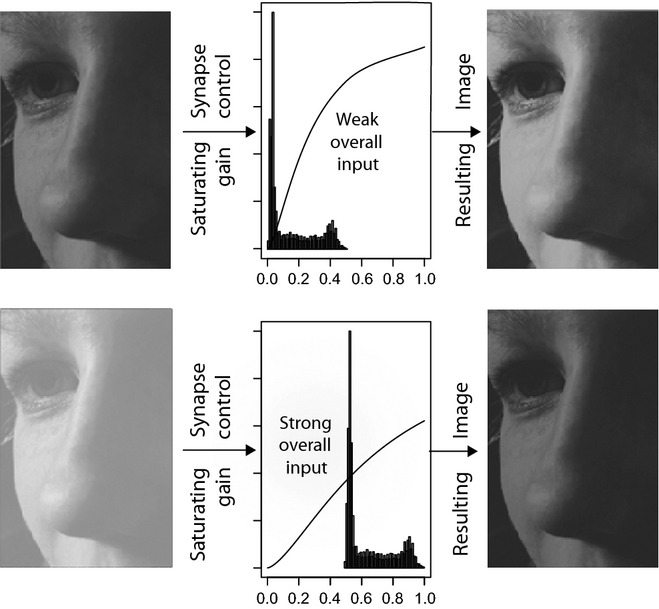
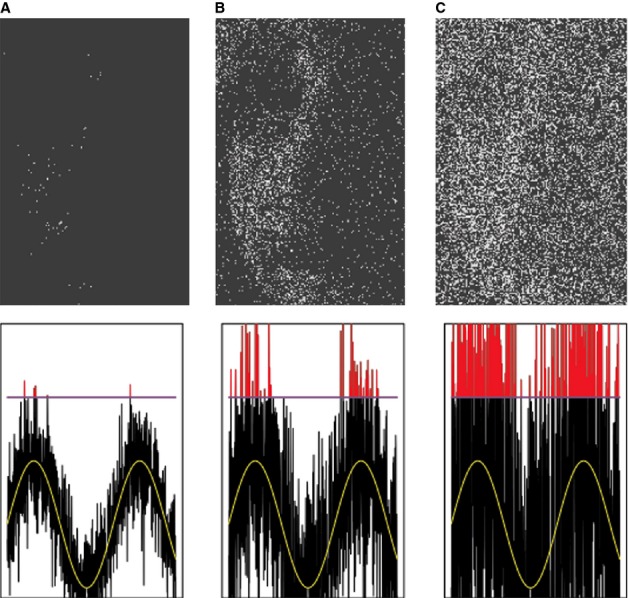
Much progress has been made recently in understanding how olfactory coding works in insect brains. Here, I propose a wiring diagram for the major steps from the first processing network (the antennal lobe) to behavioral readout. I argue that the sequence of lateral inhibition in the antennal lobe, non-linear synapses, threshold-regulating gated spring network, selective lateral inhibitory networks across glomeruli, and feedforward inhibition to the lateral protocerebrum cover most of the experimental results from different research groups and model species. I propose that the main difference between mushroom bodies and the lateral protocerebrum is not about learned vs. innate behavior. Rather, mushroom bodies perform odor identification, whereas the lateral protocerebrum performs odor evaluation (both learned and innate). I discuss the concepts of labeled line and combinatorial coding and postulate that, under restrictive experimental conditions, these networks lead to an apparent existence of 'labeled line' coding for special odors. Modulatory networks are proposed as switches between different evaluating systems in the lateral protocerebrum. A review of experimental data and theoretical conjectures both contribute to this synthesis, creating new hypotheses for future research.
Keywords: antennal lobe; mushroom bodies; neural networks; sensory coding.
© 2014 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures

Similar articles
-
The role of the Drosophila lateral horn in olfactory information processing and behavioral response.J Insect Physiol. 2017 Apr;98:29-37. doi: 10.1016/j.jinsphys.2016.11.007. Epub 2016 Nov 19. J Insect Physiol. 2017. PMID: 27871975 Review.
-
Avoidance response to CO2 in the lateral horn.PLoS Biol. 2019 Jan 17;17(1):e2006749. doi: 10.1371/journal.pbio.2006749. eCollection 2019 Jan. PLoS Biol. 2019. PMID: 30653496 Free PMC article.
-
Testing odor response stereotypy in the Drosophila mushroom body.Neuron. 2008 Sep 25;59(6):1009-23. doi: 10.1016/j.neuron.2008.07.040. Neuron. 2008. PMID: 18817738 Free PMC article.
-
Synthesis of Conserved Odor Object Representations in a Random, Divergent-Convergent Network.Neuron. 2020 Oct 28;108(2):367-381.e5. doi: 10.1016/j.neuron.2020.07.029. Epub 2020 Aug 18. Neuron. 2020. PMID: 32814018
-
Traces of Drosophila memory.Neuron. 2011 Apr 14;70(1):8-19. doi: 10.1016/j.neuron.2011.03.012. Neuron. 2011. PMID: 21482352 Free PMC article. Review.
Cited by
-
GPCR voltage dependence controls neuronal plasticity and behavior.Nat Commun. 2021 Dec 13;12(1):7252. doi: 10.1038/s41467-021-27593-x. Nat Commun. 2021. PMID: 34903750 Free PMC article.
-
The Efficacy of Some Commercially Available Insect Repellents for Aedes aegypti (Diptera: Culicidae) and Aedes albopictus (Diptera: Culicidae).J Insect Sci. 2015;15(1):140. doi: 10.1093/jisesa/iev125. J Insect Sci. 2015. PMID: 26443777 Free PMC article.
-
How bumblebees manage conflicting information seen on arrival and departure from flowers.Anim Cogn. 2025 Feb 5;28(1):11. doi: 10.1007/s10071-024-01926-x. Anim Cogn. 2025. PMID: 39909894 Free PMC article.
-
Neuroanatomical basis of sexual dimorphism in the mosquito brain.iScience. 2022 Oct 2;25(11):105255. doi: 10.1016/j.isci.2022.105255. eCollection 2022 Nov 18. iScience. 2022. PMID: 36277452 Free PMC article.
-
Olfactory responses of Drosophila are encoded in the organization of projection neurons.Elife. 2022 Sep 29;11:e77748. doi: 10.7554/eLife.77748. Elife. 2022. PMID: 36173095 Free PMC article.
References
-
- Bargmann CI. Beyond the connectome: how neuromodulators shape neural circuits. BioEssays. 2012;34:458–465. - PubMed
-
- Bargmann CI. Kaplan JM. Signal transduction in the Caenorhabditis elegans nervous system. Annu. Rev. Neurosci. 1998;21:279–308. - PubMed
-
- Barrozo RB, Gadenne C. Anton S. Switching attraction to inhibition: mating-induced reversed role of sex pheromone in an insect. J. Exp. Biol. 2010;213:2933–2939. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
