

Alveolar macrophages are essential for protection from respiratory failure and associated morbidity following influenza virus infection
- PMID: 24699679
- PMCID: PMC3974877
- DOI: 10.1371/journal.ppat.1004053
Alveolar macrophages are essential for protection from respiratory failure and associated morbidity following influenza virus infection
Abstract
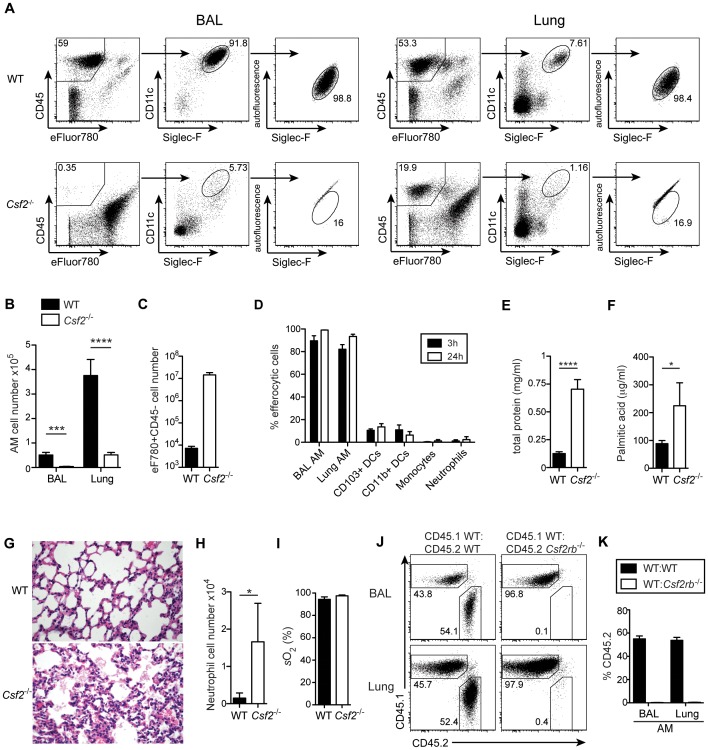
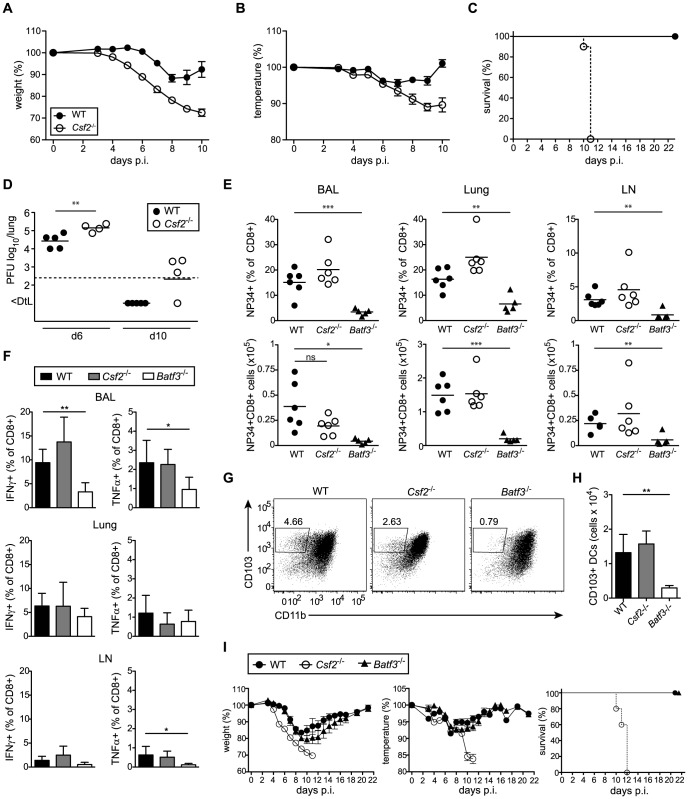
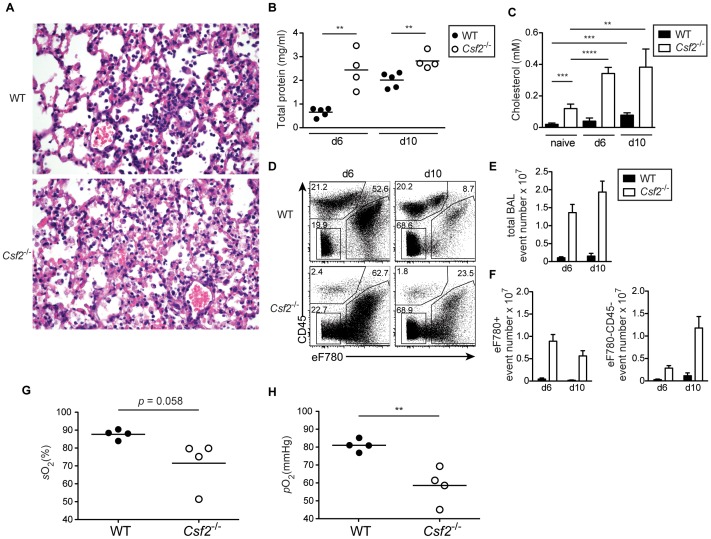
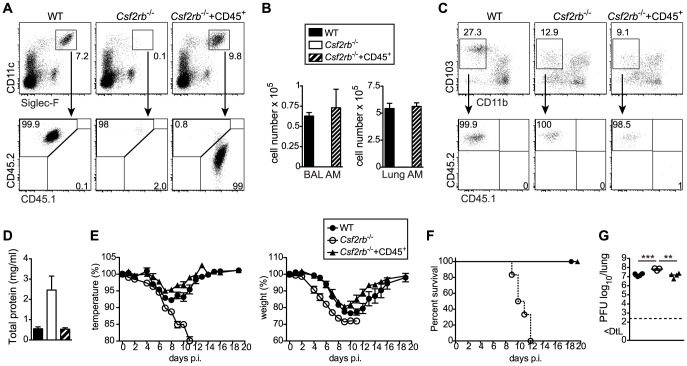
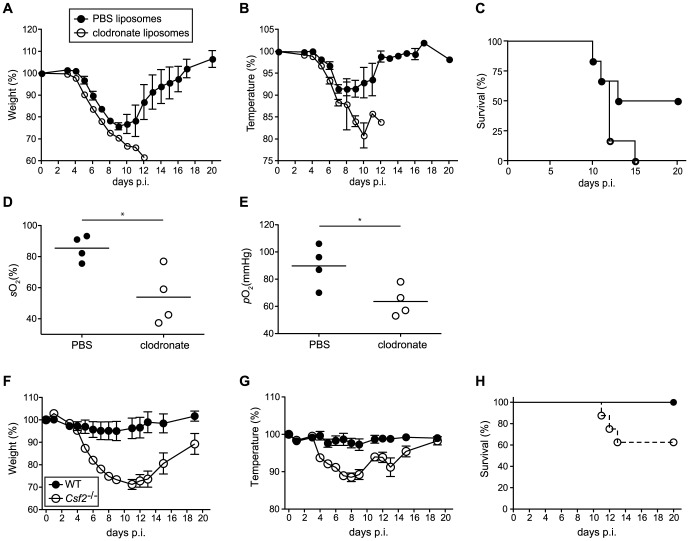
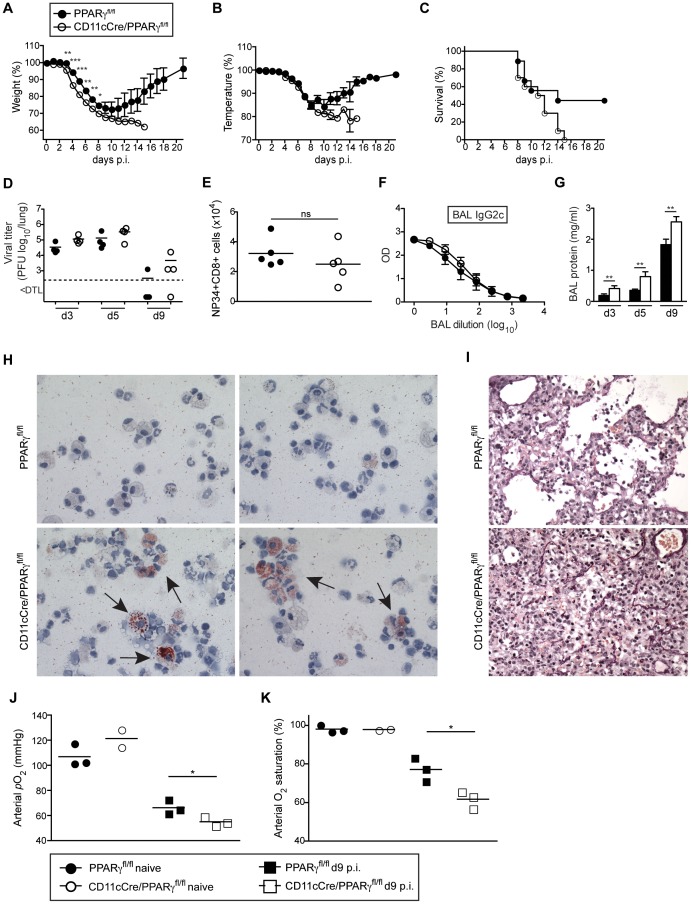
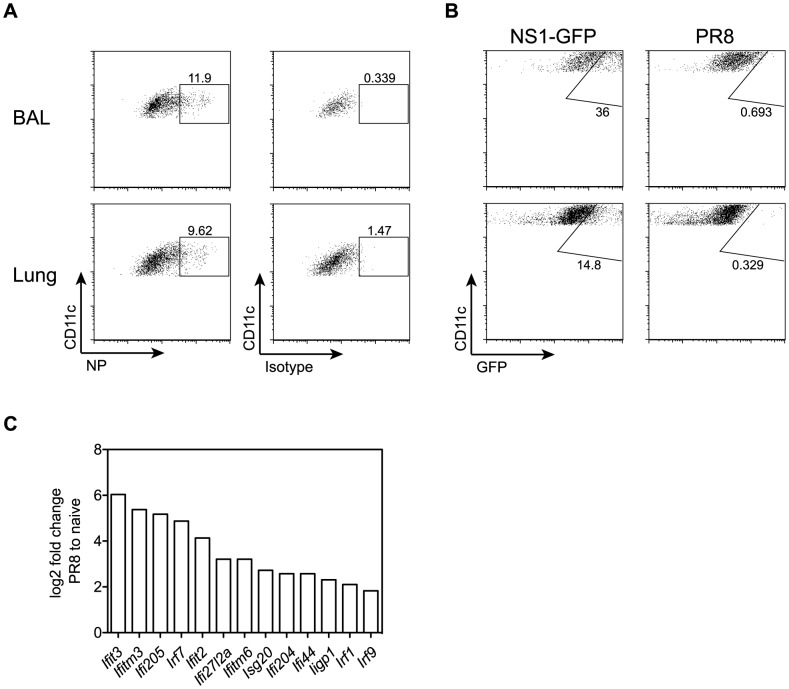
Alveolar macrophages (AM) are critical for defense against bacterial and fungal infections. However, a definitive role of AM in viral infections remains unclear. We here report that AM play a key role in survival to influenza and vaccinia virus infection by maintaining lung function and thereby protecting from asphyxiation. Absence of AM in GM-CSF-deficient (Csf2-/-) mice or selective AM depletion in wild-type mice resulted in impaired gas exchange and fatal hypoxia associated with severe morbidity to influenza virus infection, while viral clearance was affected moderately. Virus-induced morbidity was far more severe in Csf2-/- mice lacking AM, as compared to Batf3-deficient mice lacking CD8α+ and CD103+ DCs. Csf2-/- mice showed intact anti-viral CD8+ T cell responses despite slightly impaired CD103+ DC development. Importantly, selective reconstitution of AM development in Csf2rb-/- mice by neonatal transfer of wild-type AM progenitors prevented severe morbidity and mortality, demonstrating that absence of AM alone is responsible for disease severity in mice lacking GM-CSF or its receptor. In addition, CD11c-Cre/Ppargfl/fl mice with a defect in AM but normal adaptive immunity showed increased morbidity and lung failure to influenza virus. Taken together, our results suggest a superior role of AM compared to CD103+ DCs in protection from acute influenza and vaccinia virus infection-induced morbidity and mortality.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Trapnell BC, Whitsett JA (2002) Gm-CSF regulates pulmonary surfactant homeostasis and alveolar macrophage-mediated innate host defense. Annual review of physiology 64: 775–802. - PubMed
-
- Trapnell BC, Whitsett JA, Nakata K (2003) Pulmonary alveolar proteinosis. The New England journal of medicine 349: 2527–2539. - PubMed
-
- Seymour JF, Presneill JJ (2002) Pulmonary alveolar proteinosis: progress in the first 44 years. American Journal of Respiratory and Critical Care Medicine 166: 215–235. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
