

The pathogenic mechanism of the Mycobacterium ulcerans virulence factor, mycolactone, depends on blockade of protein translocation into the ER
- PMID: 24699819
- PMCID: PMC3974873
- DOI: 10.1371/journal.ppat.1004061
The pathogenic mechanism of the Mycobacterium ulcerans virulence factor, mycolactone, depends on blockade of protein translocation into the ER
Abstract
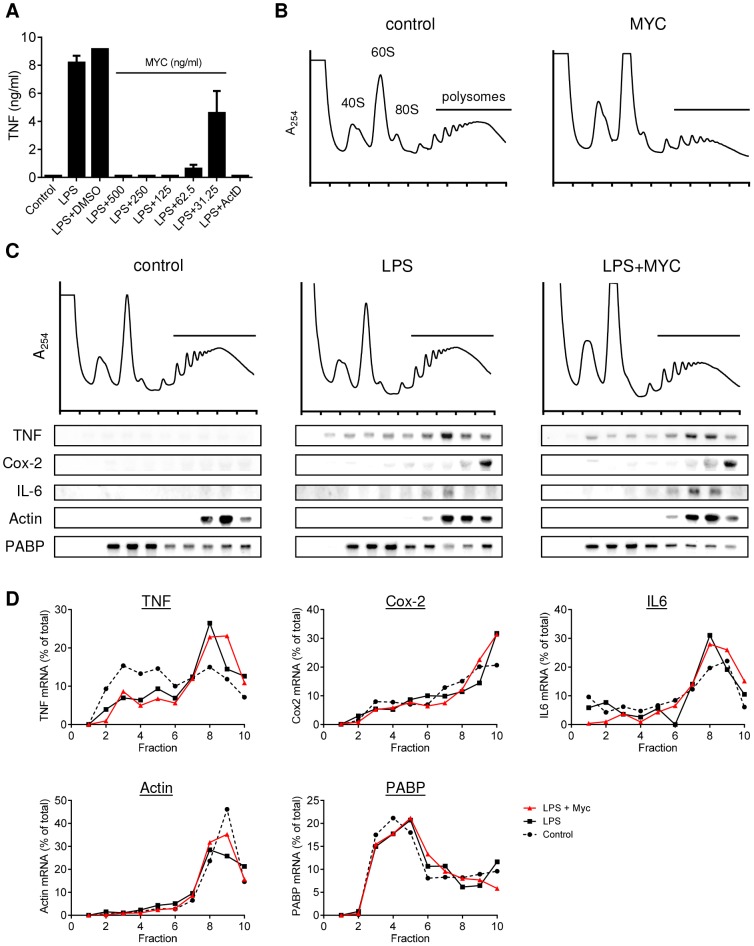
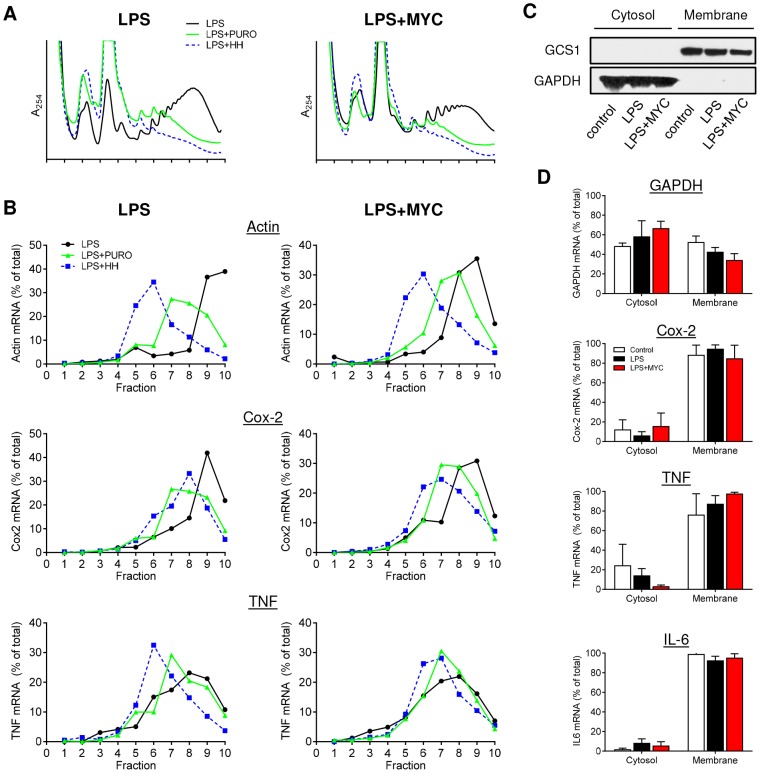
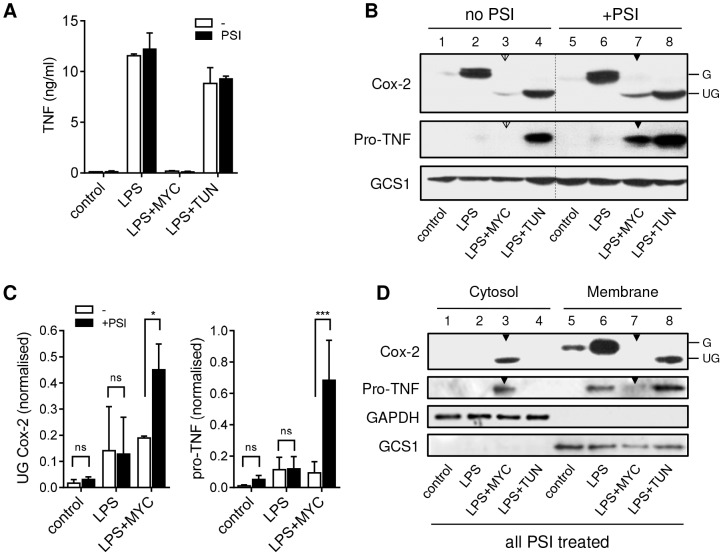
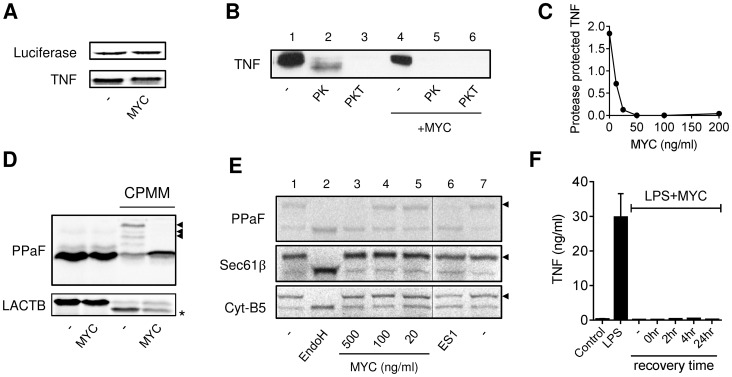
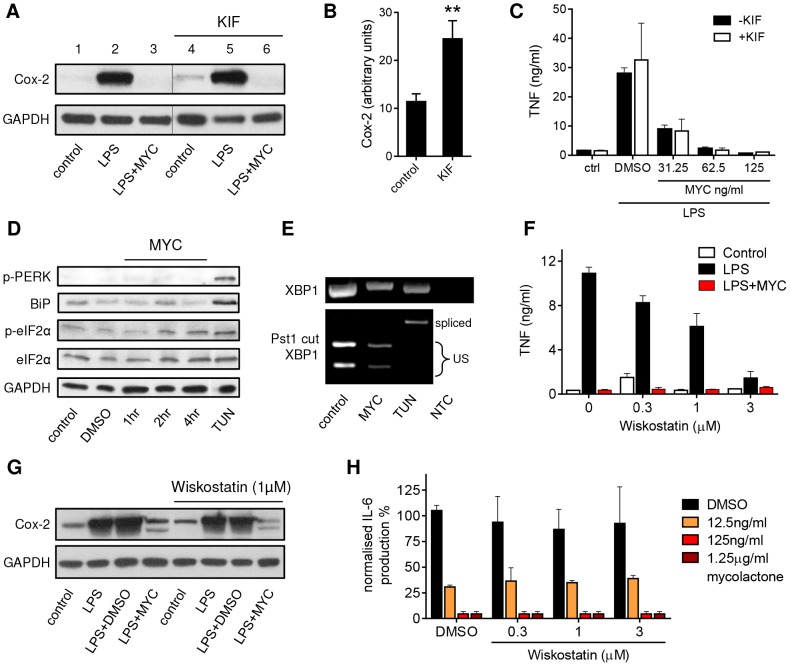
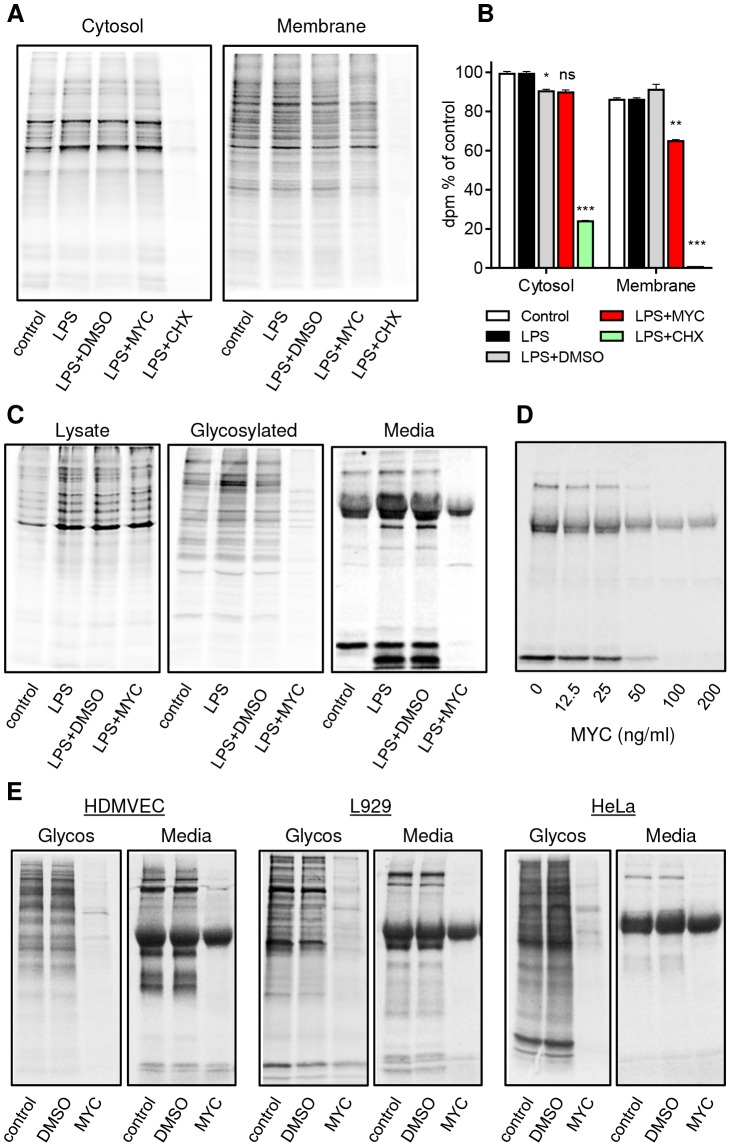
Infection with Mycobacterium ulcerans is characterised by tissue necrosis and immunosuppression due to mycolactone, the necessary and sufficient virulence factor for Buruli ulcer disease pathology. Many of its effects are known to involve down-regulation of specific proteins implicated in important cellular processes, such as immune responses and cell adhesion. We have previously shown mycolactone completely blocks the production of LPS-dependent proinflammatory mediators post-transcriptionally. Using polysome profiling we now demonstrate conclusively that mycolactone does not prevent translation of TNF, IL-6 and Cox-2 mRNAs in macrophages. Instead, it inhibits the production of these, along with nearly all other (induced and constitutive) proteins that transit through the ER. This is due to a blockade of protein translocation and subsequent degradation of aberrantly located protein. Several lines of evidence support this transformative explanation of mycolactone function. First, cellular TNF and Cox-2 can be once more detected if the action of the 26S proteasome is inhibited concurrently. Second, restored protein is found in the cytosol, indicating an inability to translocate. Third, in vitro translation assays show mycolactone prevents the translocation of TNF and other proteins into the ER. This is specific as the insertion of tail-anchored proteins into the ER is unaffected showing that the ER remains structurally intact. Fourth, metabolic labelling reveals a near-complete loss of glycosylated and secreted proteins from treated cells, whereas cytosolic proteins are unaffected. Notably, the profound lack of glycosylated and secreted protein production is apparent in a range of different disease-relevant cell types. These studies provide a new mechanism underlying mycolactone's observed pathological activities both in vitro and in vivo. Mycolactone-dependent inhibition of protein translocation into the ER not only explains the deficit of innate cytokines, but also the loss of membrane receptors, adhesion molecules and T-cell cytokines that drive the aetiology of Buruli ulcer.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- George KM, Chatterjee D, Gunawardana G, Welty D, Hayman J, et al. (1999) Mycolactone: a polyketide toxin from Mycobacterium ulcerans required for virulence. Science 283: 854–857. - PubMed
-
- Demangel C, Stinear TP, Cole ST (2009) Buruli ulcer: reductive evolution enhances pathogenicity of Mycobacterium ulcerans. Nat Rev Microbiol 7: 50–60. - PubMed
-
- Hall BS, Simmonds RE (2013) Pleiotropic molecular effects of the Mycobacterium ulcerans virulence factor mycolactone underlying the cell death and immunosuppression seen in Buruli ulcer. Biochem Soc Trans 42: 177–183. - PubMed
-
- Simmonds RE, Lali FV, Smallie T, Small PL, Foxwell BM (2009) Mycolactone inhibits monocyte cytokine production by a posttranscriptional mechanism. J Immunol 182: 2194–2202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
