

ATR checkpoint kinase and CRL1βTRCP collaborate to degrade ASF1a and thus repress genes overlapping with clusters of stalled replication forks
- PMID: 24700029
- PMCID: PMC4003279
- DOI: 10.1101/gad.239194.114
ATR checkpoint kinase and CRL1βTRCP collaborate to degrade ASF1a and thus repress genes overlapping with clusters of stalled replication forks
Abstract
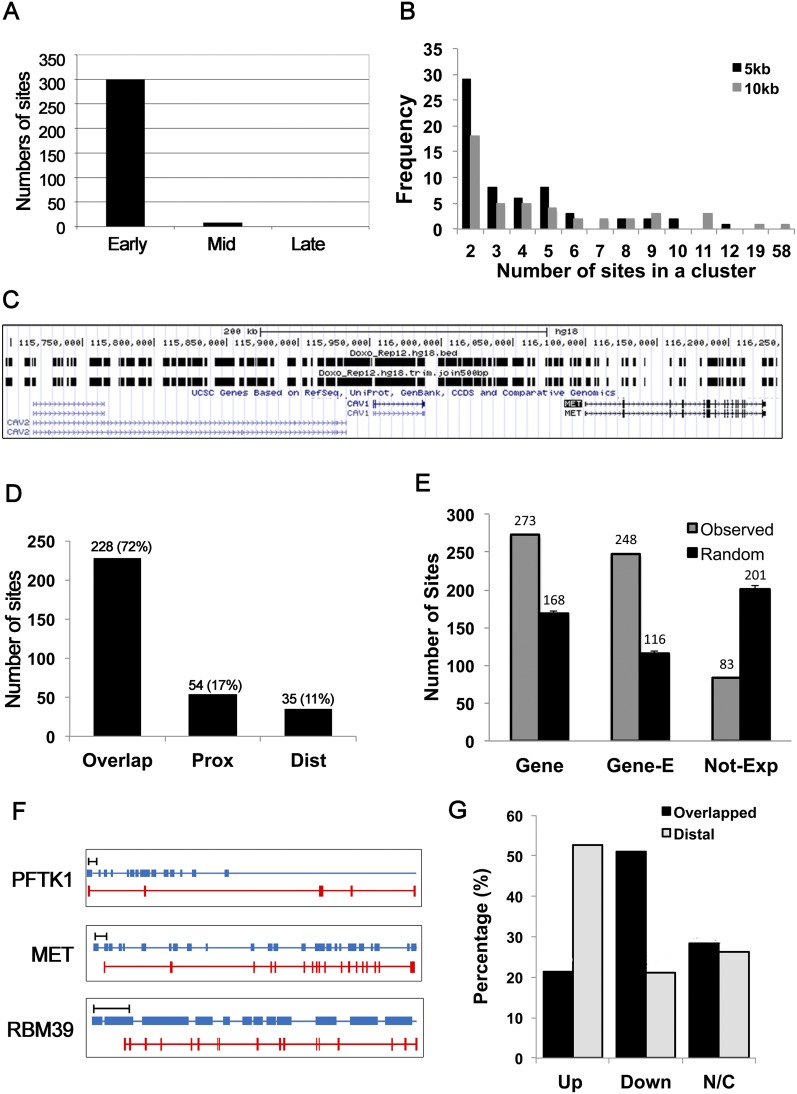
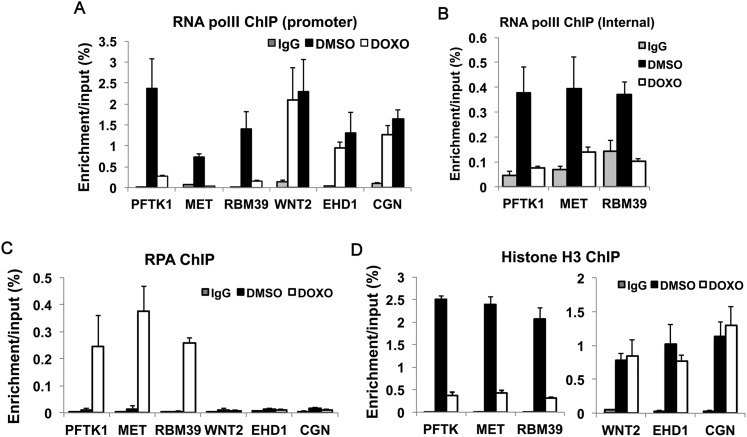
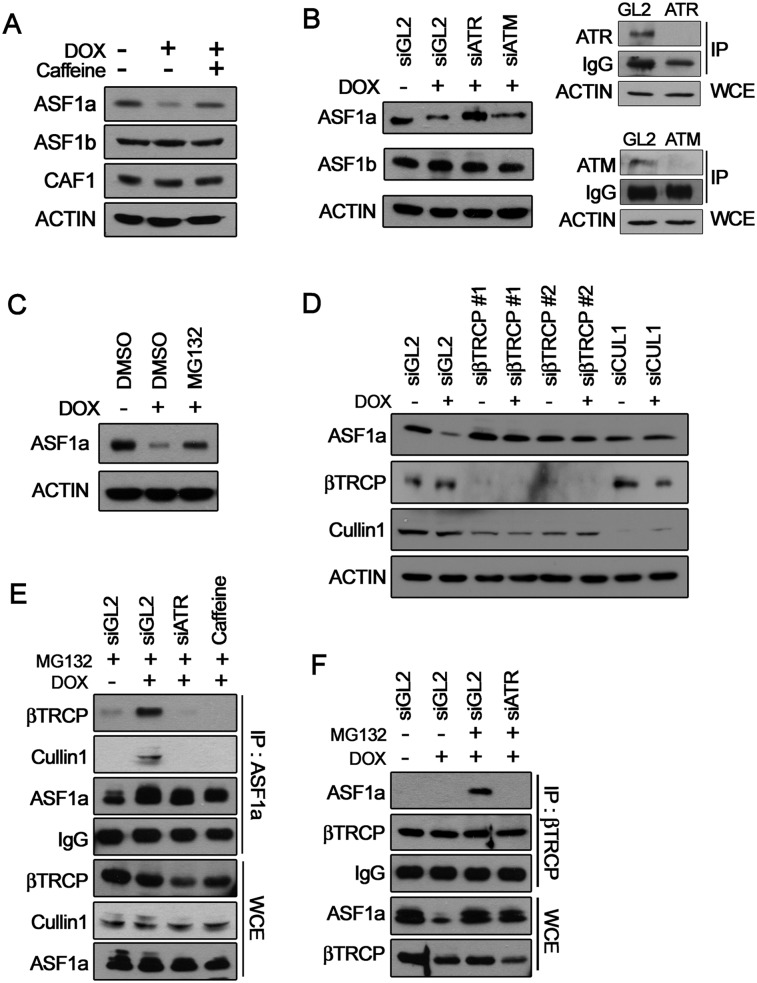
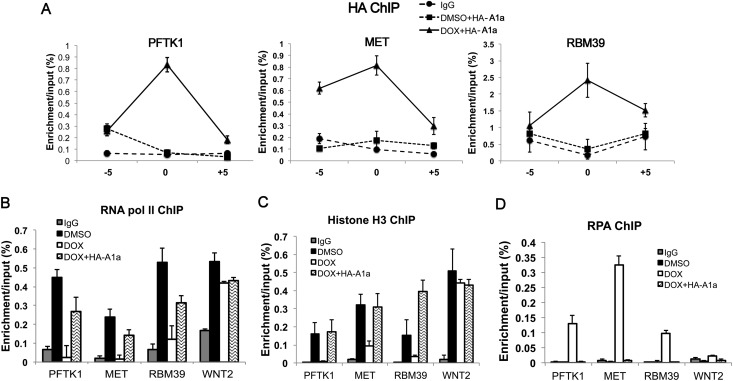
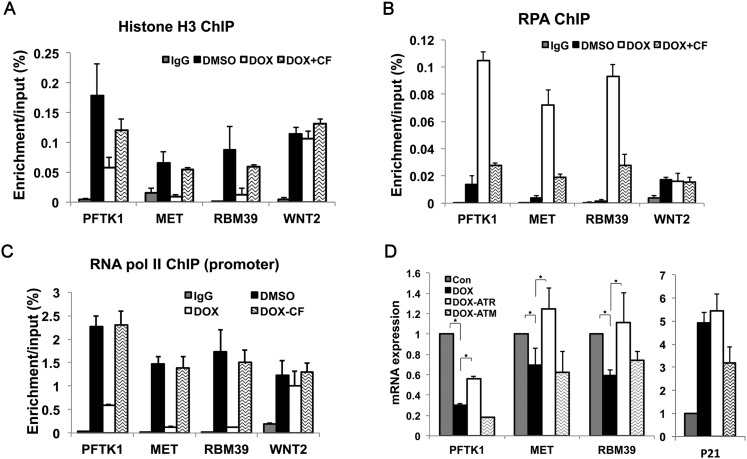
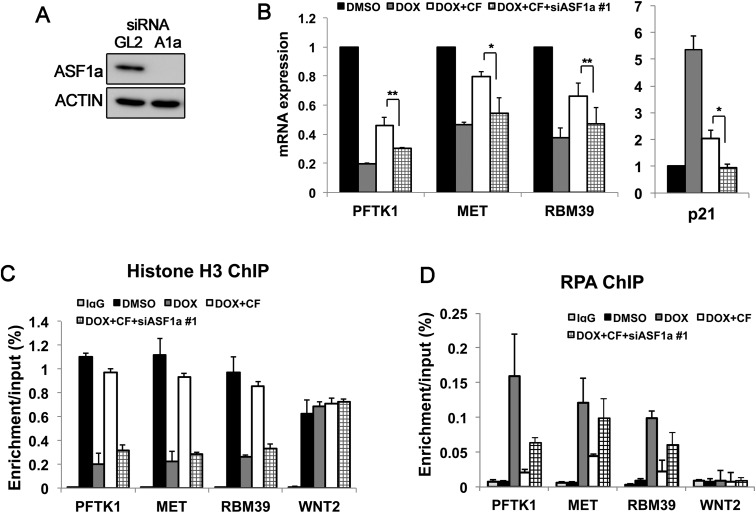
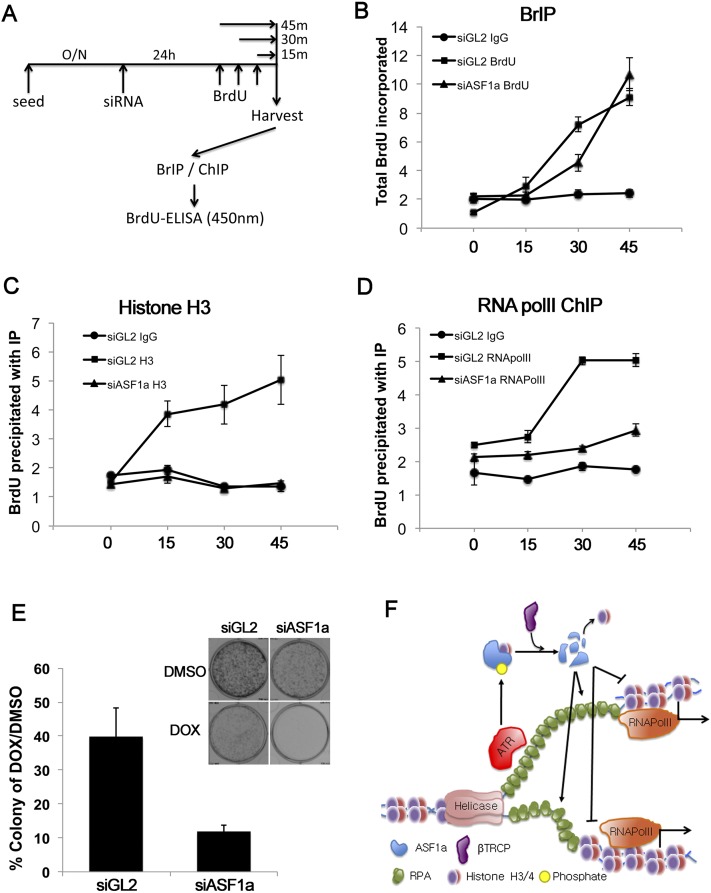
Many agents used for chemotherapy, such as doxorubicin, interfere with DNA replication, but the effect of this interference on transcription is largely unknown. Here we show that doxorubicin induces the firing of dense clusters of neoreplication origins that lead to clusters of stalled replication forks in gene-rich parts of the genome, particularly on expressed genes. Genes that overlap with these clusters of stalled forks are actively dechromatinized, unwound, and repressed by an ATR-dependent checkpoint pathway. The ATR checkpoint pathway causes a histone chaperone normally associated with the replication fork, ASF1a, to degrade through a CRL1(βTRCP)-dependent ubiquitination/proteasome pathway, leading to the localized dechromatinization and gene repression. Therefore, a globally active checkpoint pathway interacts with local clusters of stalled forks to specifically repress genes in the vicinity of the stalled forks, providing a new mechanism of action of chemotherapy drugs like doxorubicin. Finally, ASF1a-depleted cancer cells are more sensitive to doxorubicin, suggesting that the 7%-10% of prostate adenocarcinomas and adenoid cystic carcinomas reported to have homozygous deletion or significant underexpression of ASF1a should be tested for high sensitivity to doxorubicin.
Keywords: ASF1; ATR; CRL1βTRCP; S-phase checkpoint; doxorubicin; stalled forks; transcription repression.
Figures
References
-
- Barr SM, Leung CG, Chang EE, Cimprich KA 2003. ATR kinase activity regulates the intranuclear translocation of ATR and RPA following ionizing radiation. Curr Biol 13: 1047–1051 - PubMed
-
- Bell SP, Dutta A 2002. DNA replication in eukaryotic cells. Annu Rev Biochem 71: 333–374 - PubMed
-
- Bodley A, Liu LF, Israel M, Seshadri R, Koseki Y, Giuliani FC, Kirschenbaum S, Silber R, Potmesil M 1989. DNA topoisomerase II-mediated interaction of doxorubicin and daunorubicin congeners with DNA. Cancer Res 49: 5969–5978 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous