

A RAD-tag genetic map for the platyfish (Xiphophorus maculatus) reveals mechanisms of karyotype evolution among teleost fish
- PMID: 24700104
- PMCID: PMC4063920
- DOI: 10.1534/genetics.114.164293
A RAD-tag genetic map for the platyfish (Xiphophorus maculatus) reveals mechanisms of karyotype evolution among teleost fish
Abstract
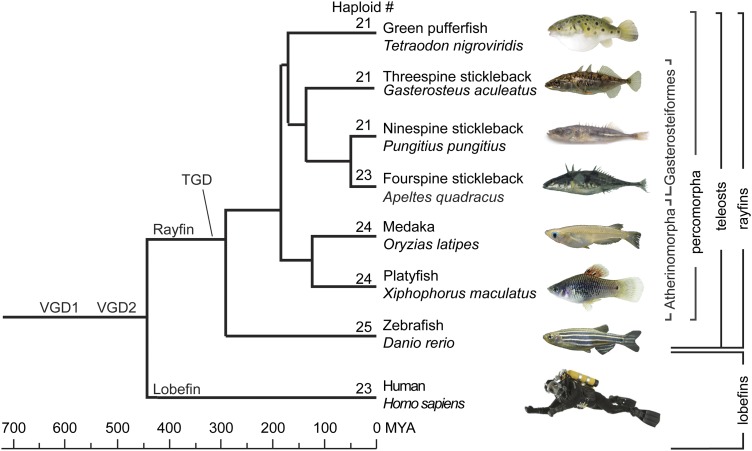
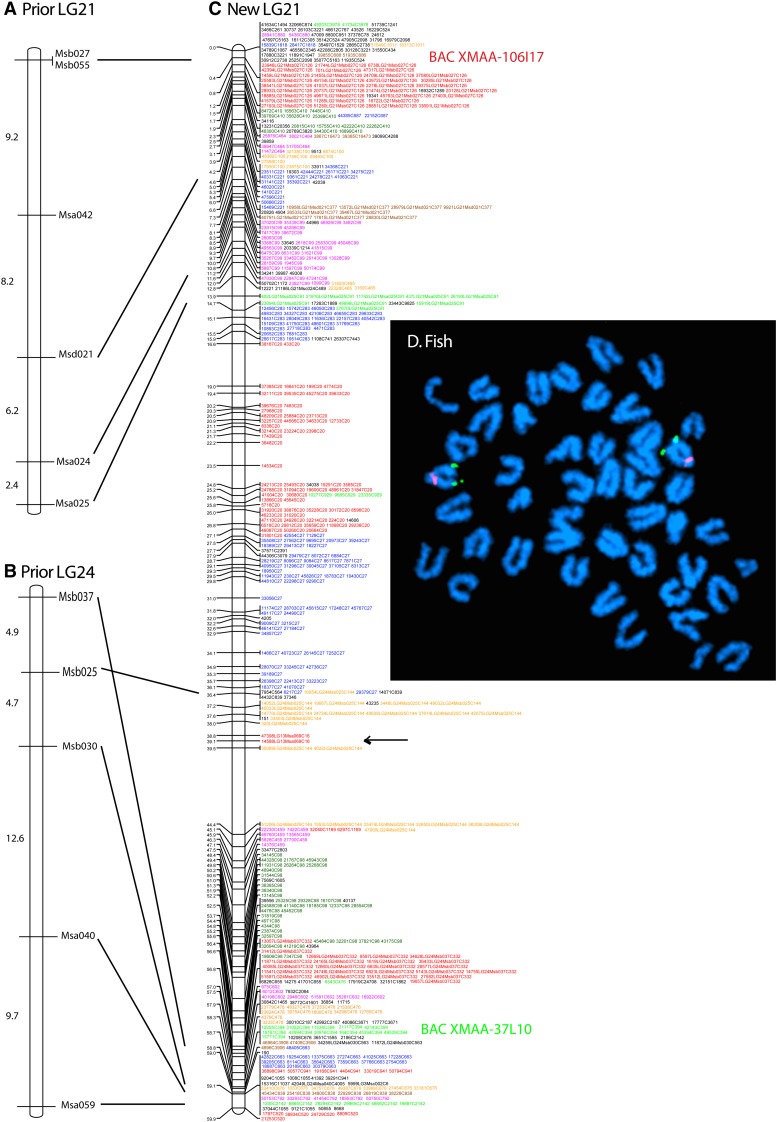
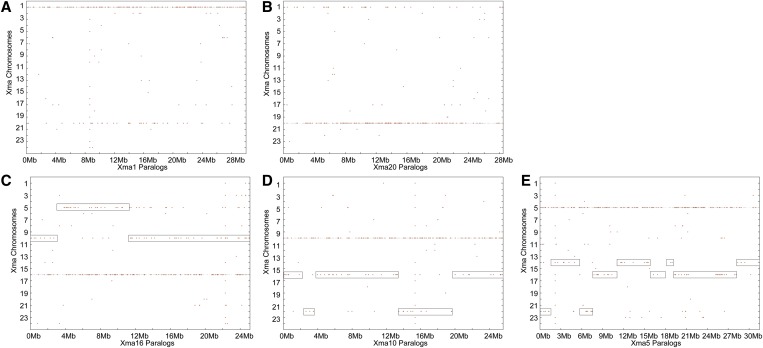
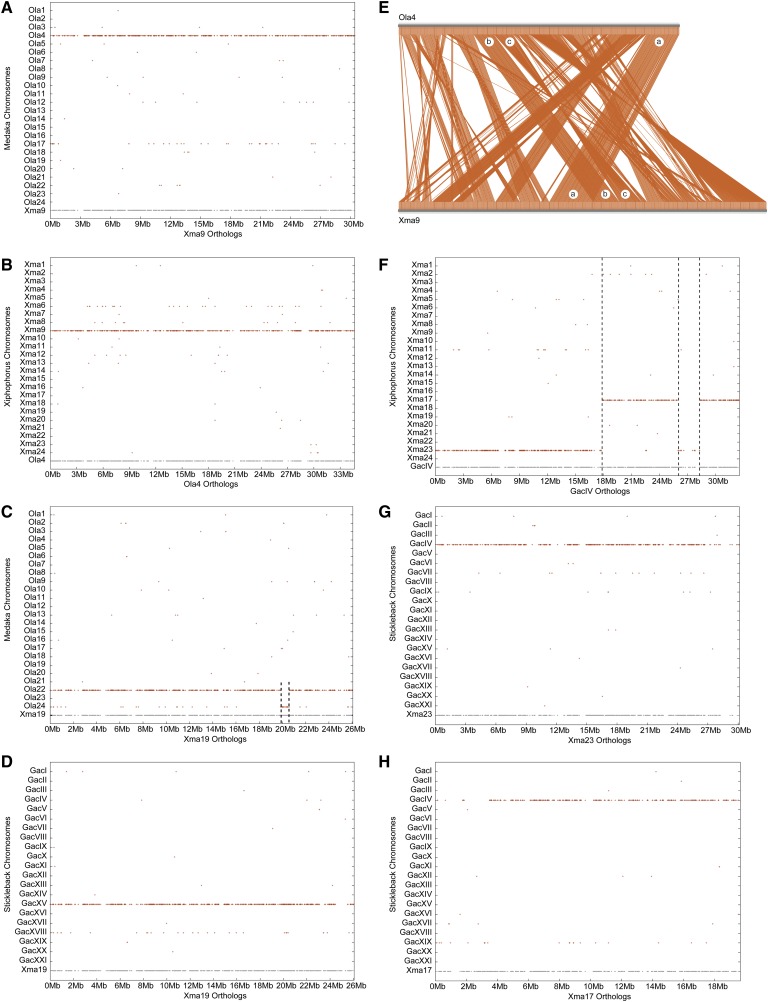
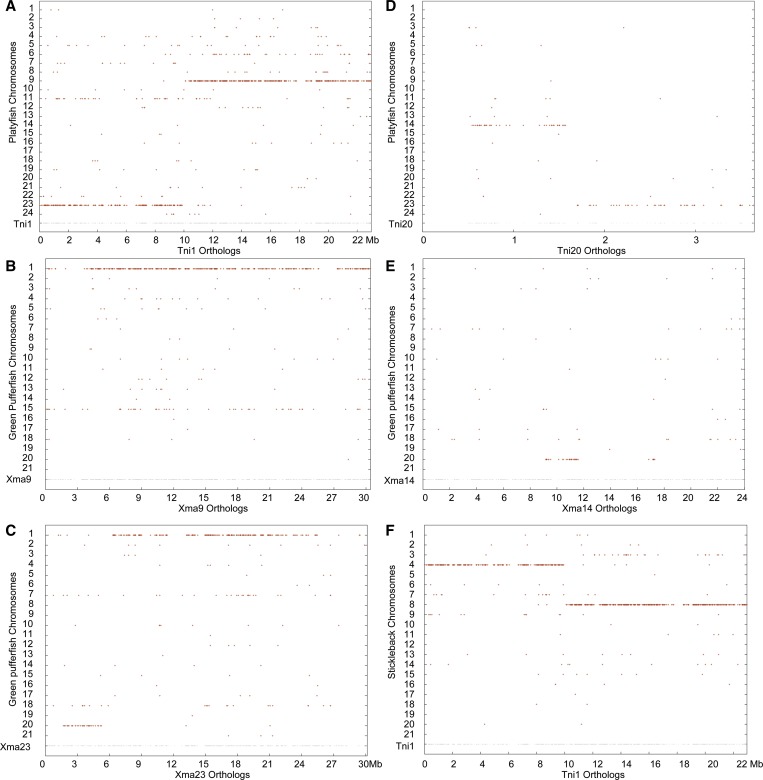
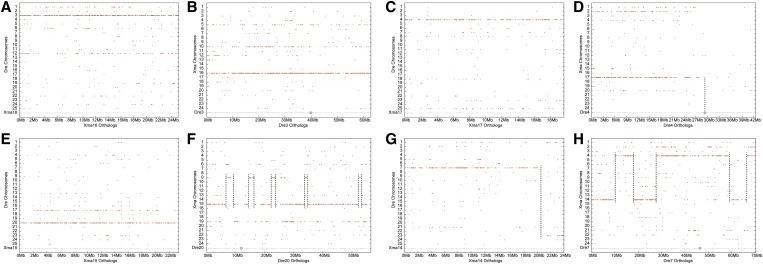
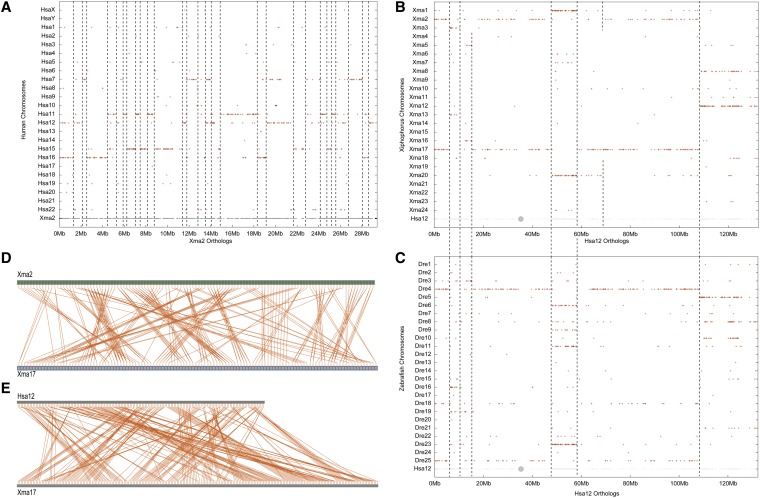
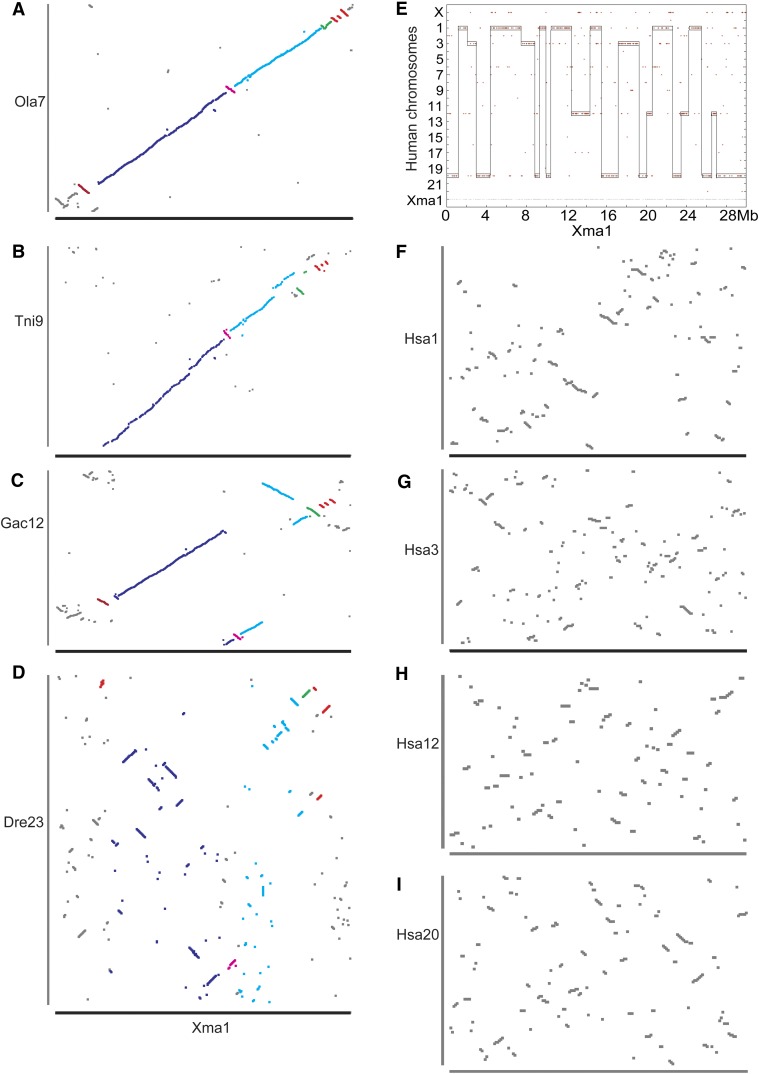
Mammalian genomes can vary substantially in haploid chromosome number even within a small taxon (e.g., 3-40 among deer alone); in contrast, teleost fish genomes are stable (24-25 in 58% of teleosts), but we do not yet understand the mechanisms that account for differences in karyotype stability. Among perciform teleosts, platyfish (Xiphophorus maculatus) and medaka (Oryzias latipes) both have 24 chromosome pairs, but threespine stickleback (Gasterosteus aculeatus) and green pufferfish (Tetraodon nigroviridis) have just 21 pairs. To understand the evolution of teleost genomes, we made a platyfish meiotic map containing 16,114 mapped markers scored on 267 backcross fish. We tiled genomic contigs along the map to create chromosome-length genome assemblies. Genome-wide comparisons of conserved synteny showed that platyfish and medaka karyotypes remained remarkably similar with few interchromosomal translocations but with numerous intrachromosomal rearrangements (transpositions and inversions) since their lineages diverged ∼120 million years ago. Comparative genomics with platyfish shows how reduced chromosome numbers in stickleback and green pufferfish arose by fusion of pairs of ancestral chromosomes after their lineages diverged from platyfish ∼195 million years ago. Zebrafish and human genomes provide outgroups to root observed changes. These studies identify likely genome assembly errors, characterize chromosome fusion events, distinguish lineage-independent chromosome fusions, show that the teleost genome duplication does not appear to have accelerated the rate of translocations, and reveal the stability of syntenies and gene orders in teleost chromosomes over hundreds of millions of years.
Keywords: conserved synteny; genome evolution; medaka; stickleback; swordtail; zebrafish.
Copyright © 2014 by the Genetics Society of America.
Figures
References
-
- Amores A., Force A., Yan Y. L., Joly L., Amemiya C., et al. , 1998. Zebrafish hox clusters and vertebrate genome evolution. Science 282: 1711–1714 - PubMed
-
- Braasch I., Postlethwait J. H., 2012. Polyploidy in fish and the teleost genome duplication, pp. 341–382 in Polyploidy and Genome Evolution, edited by Soltis P., Soltis D. Springer, New York
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
