

aPKCλ maintains the integrity of the glomerular slit diaphragm through trafficking of nephrin to the cell surface
- PMID: 24700503
- PMCID: PMC4112437
- DOI: 10.1093/jb/mvu022
aPKCλ maintains the integrity of the glomerular slit diaphragm through trafficking of nephrin to the cell surface
Abstract
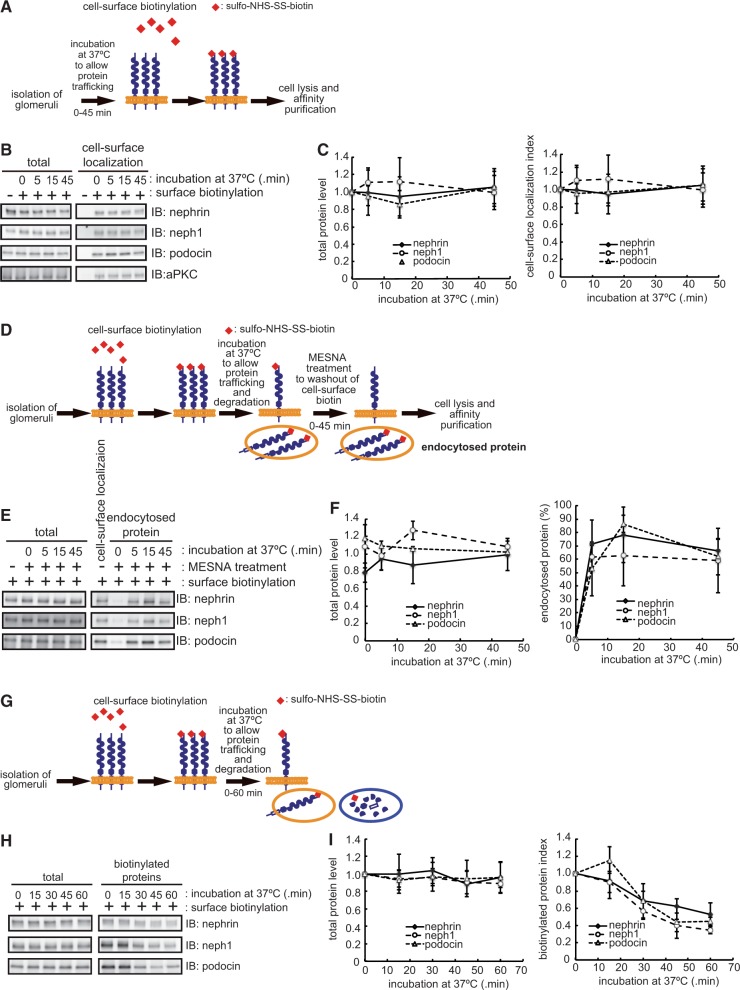
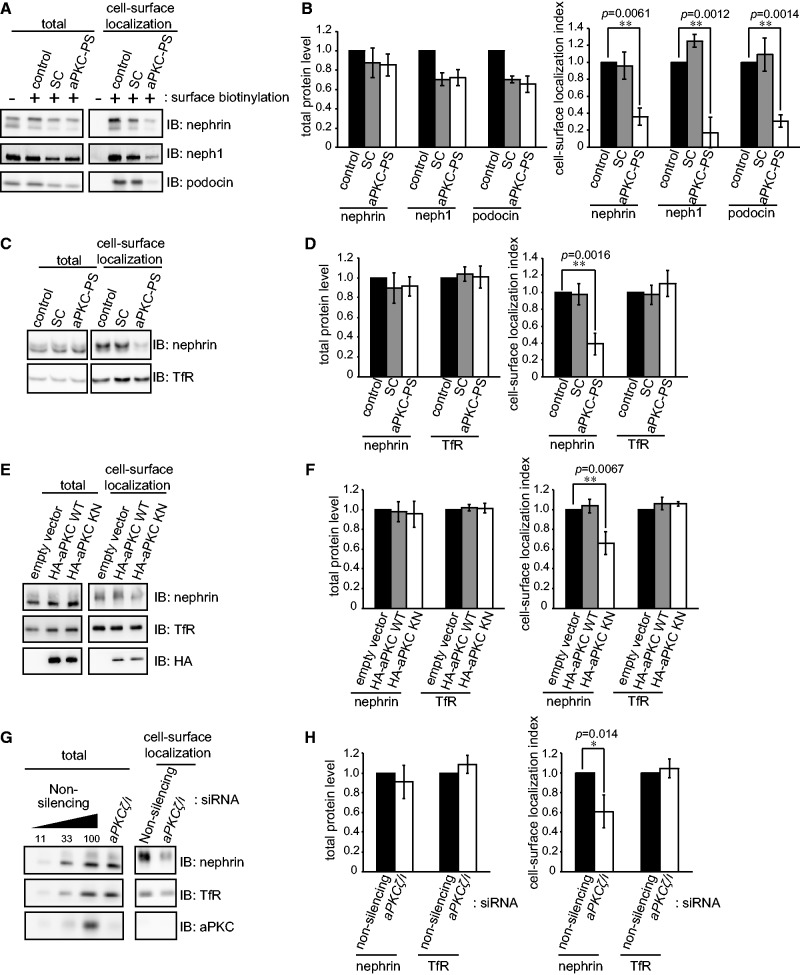
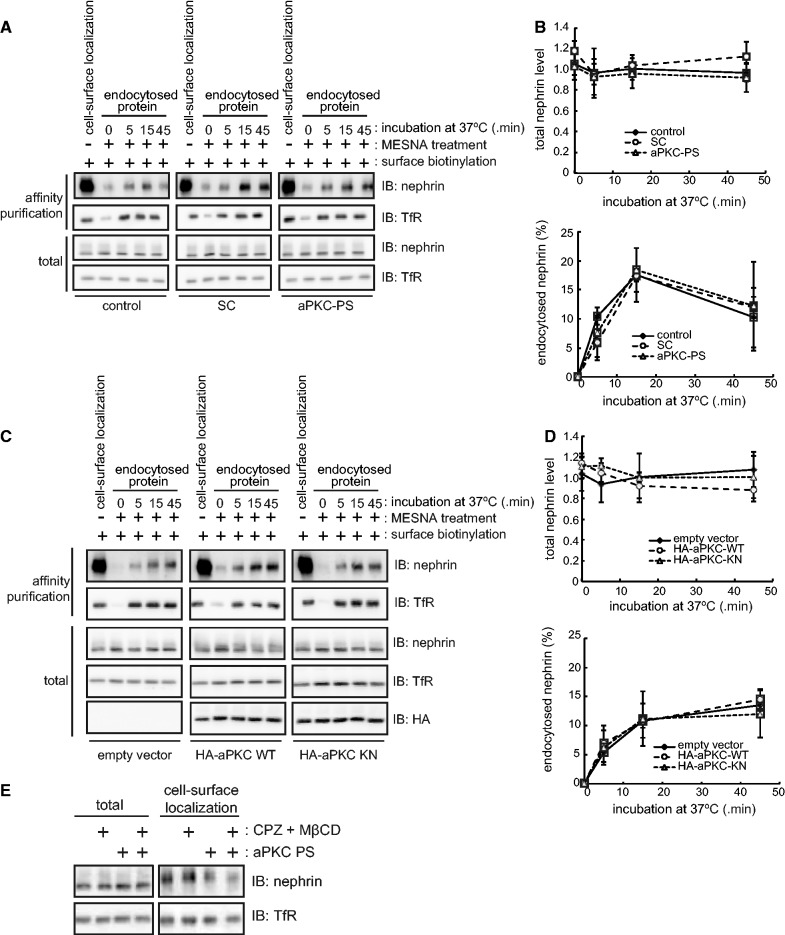
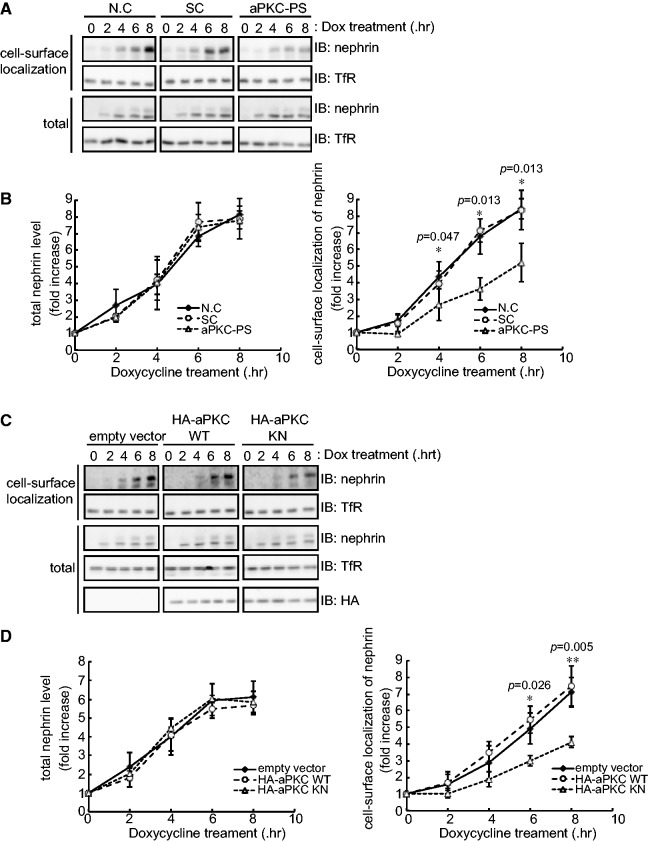
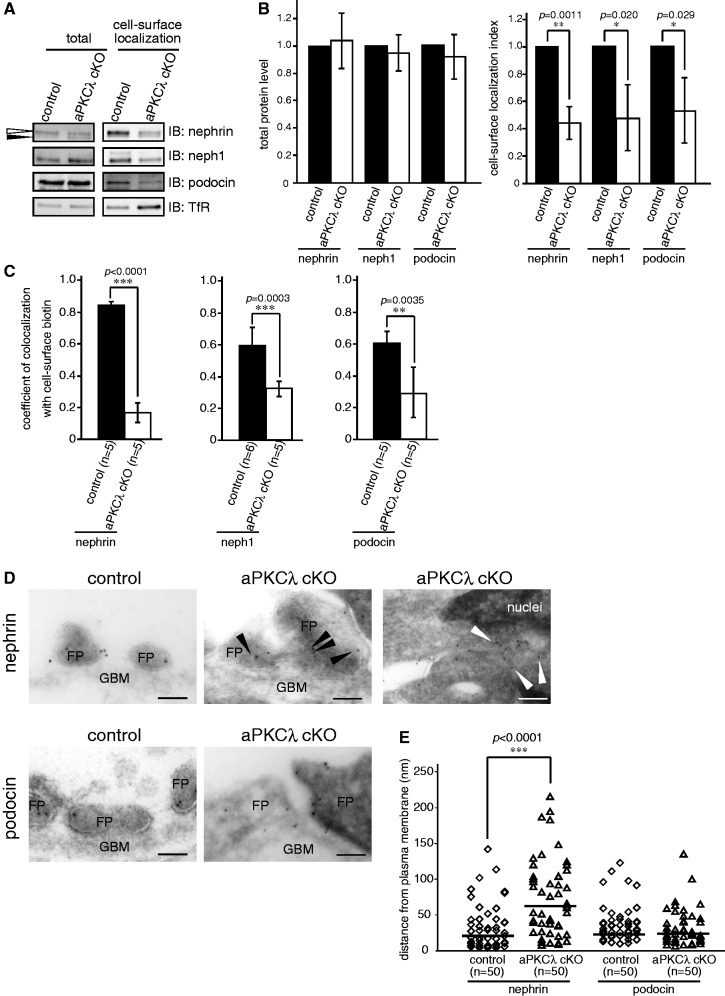
The slit diaphragm (SD), the specialized intercellular junction between renal glomerular epithelial cells (podocytes), provides a selective-filtration barrier in renal glomeruli. Dysfunction of the SD results in glomerular diseases that are characterized by disappearance of SD components, such as nephrin, from the cell surface. Although the importance of endocytosis and degradation of SD components for the maintenance of SD integrity has been suggested, the dynamic nature of the turnover of intact cell-surface SD components remained unclear. Using isolated rat glomeruli we show that the turnover rates of cell-surface SD components are relatively high; they almost completely disappear from the cell surface within minutes. The exocytosis, but not endocytosis, of heterologously expressed nephrin requires the kinase activity of the cell polarity regulator atypical protein kinase C (aPKC). Consistently, we demonstrate that podocyte-specific deletion of aPKCλ resulted in a decrease of cell-surface localization of SD components, causing massive proteinuria. In conclusion, the regulation of SD turnover by aPKC is crucial for the maintenance of SD integrity and defects in aPKC signalling can lead to proteinuria. These findings not only reveal the pivotal importance of the dynamic turnover of cell-surface SD components but also suggest a novel pathophysiological basis in glomerular disease.
Keywords: aPKC; cell-surface localization; glomerular disease; nephrin; slit diaphragm.
© The Authors 2014. Published by Oxford University Press on behalf of the Japanese Biochemical Society.
Figures
Similar articles
-
An essential role of the universal polarity protein, aPKClambda, on the maintenance of podocyte slit diaphragms.PLoS One. 2009;4(1):e4194. doi: 10.1371/journal.pone.0004194. Epub 2009 Jan 14. PLoS One. 2009. PMID: 19142224 Free PMC article.
-
Loss of podocyte aPKClambda/iota causes polarity defects and nephrotic syndrome.J Am Soc Nephrol. 2009 Apr;20(4):798-806. doi: 10.1681/ASN.2008080871. Epub 2009 Mar 11. J Am Soc Nephrol. 2009. PMID: 19279126 Free PMC article.
-
Disruption of the exocyst induces podocyte loss and dysfunction.J Biol Chem. 2019 Jun 28;294(26):10104-10119. doi: 10.1074/jbc.RA119.008362. Epub 2019 May 9. J Biol Chem. 2019. PMID: 31073028 Free PMC article.
-
Slit diaphragm dysfunction in proteinuric states: identification of novel therapeutic targets for nephrotic syndrome.Clin Exp Nephrol. 2009 Aug;13(4):275-280. doi: 10.1007/s10157-009-0162-x. Epub 2009 Mar 7. Clin Exp Nephrol. 2009. PMID: 19266252 Review.
-
Endocytic Trafficking at the Mature Podocyte Slit Diaphragm.Front Pediatr. 2017 Feb 24;5:32. doi: 10.3389/fped.2017.00032. eCollection 2017. Front Pediatr. 2017. PMID: 28286744 Free PMC article. Review.
Cited by
-
An acytokinetic cell division creates PIP2-enriched membrane asymmetries leading to slit diaphragm assembly in Drosophila nephrocytes.Development. 2023 Sep 15;150(18):dev201708. doi: 10.1242/dev.201708. Epub 2023 Sep 18. Development. 2023. PMID: 37681291 Free PMC article.
-
Mature induced-pluripotent-stem-cell-derived human podocytes reconstitute kidney glomerular-capillary-wall function on a chip.Nat Biomed Eng. 2017;1:0069. doi: 10.1038/s41551-017-0069. Epub 2017 May 10. Nat Biomed Eng. 2017. PMID: 29038743 Free PMC article.
-
Recent advances of animal model of focal segmental glomerulosclerosis.Clin Exp Nephrol. 2018 Aug;22(4):752-763. doi: 10.1007/s10157-018-1552-8. Epub 2018 Mar 20. Clin Exp Nephrol. 2018. PMID: 29556761 Review.
-
Defects of CRB2 cause steroid-resistant nephrotic syndrome.Am J Hum Genet. 2015 Jan 8;96(1):153-61. doi: 10.1016/j.ajhg.2014.11.014. Epub 2014 Dec 31. Am J Hum Genet. 2015. PMID: 25557779 Free PMC article.
-
Role of protein kinase C in podocytes and development of glomerular damage in diabetic nephropathy.Front Endocrinol (Lausanne). 2014 Nov 5;5:179. doi: 10.3389/fendo.2014.00179. eCollection 2014. Front Endocrinol (Lausanne). 2014. PMID: 25414693 Free PMC article. Review.
References
-
- Tryggvason K, Patrakka J, Wartiovaara J. Hereditary proteinuria syndromes and mechanisms of proteinuria. N. Engl. J. Med. 2006;354:1387–1401. - PubMed
-
- Welsh GI, Saleem MA. Nephrin-signature molecule of the glomerular podocyte? J. Pathol. 2010;220:328–337. - PubMed
-
- Kestila M, Lenkkeri U, Mannikko M, Lamerdin J, McCready P, Putaala H, Ruotsalainen V, Morita T, Nissinen M, Herva R, Kashtan CE, Peltonen L, Holmberg C, Olsen A, Tryggvason K. Positionally cloned gene for a novel glomerular protein—nephrin—is mutated in congenital nephrotic syndrome. Mol. Cell. 1998;1:575–582. - PubMed
-
- Putaala H, Soininen R, Kilpelainen P, Wartiovaara J, Tryggvason K. The murine nephrin gene is specifically expressed in kidney, brain and pancreas: inactivation of the gene leads to massive proteinuria and neonatal death. Hum. Mol. Genet. 2001;10:1–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
