

PTK7-Src signaling at epithelial cell contacts mediates spatial organization of actomyosin and planar cell polarity
- PMID: 24703874
- PMCID: PMC4086913
- DOI: 10.1016/j.devcel.2014.02.008
PTK7-Src signaling at epithelial cell contacts mediates spatial organization of actomyosin and planar cell polarity
Abstract
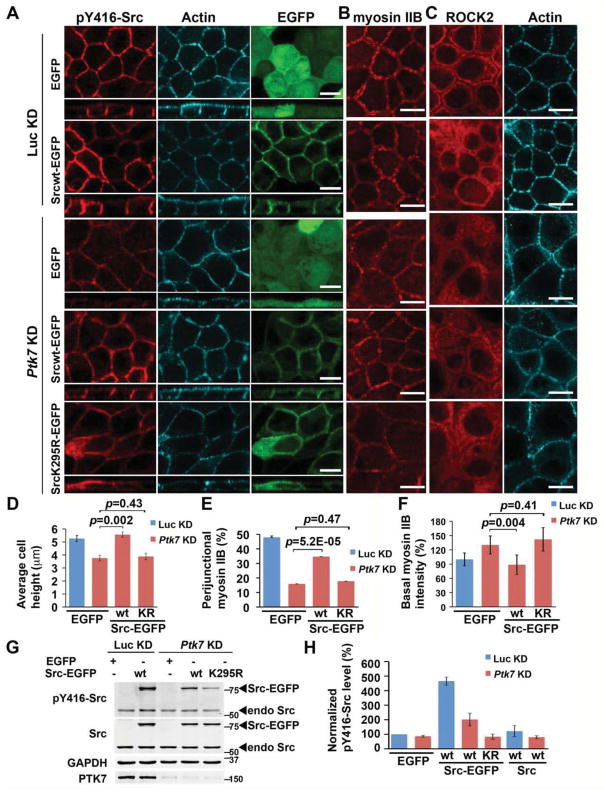
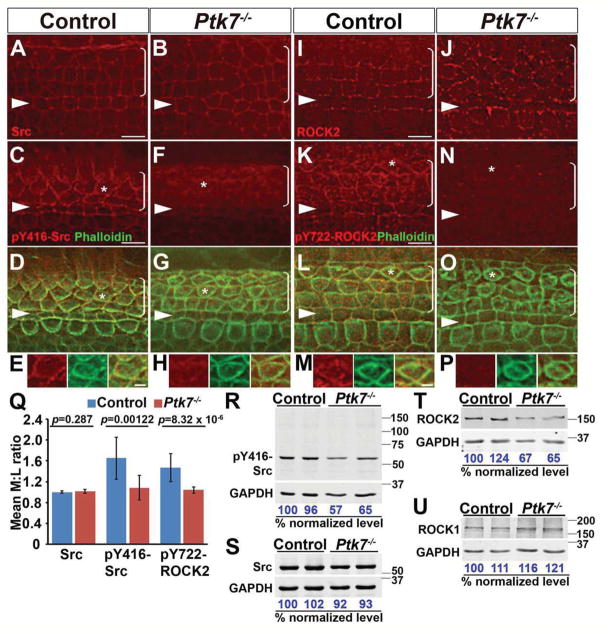
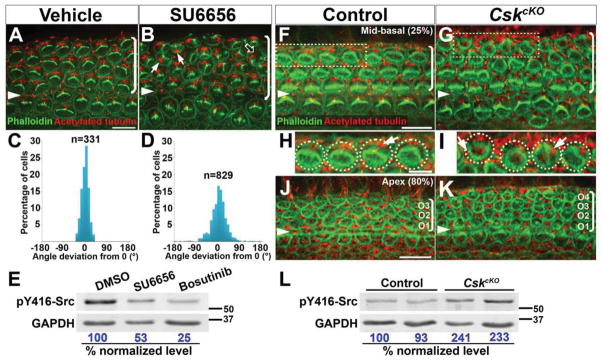
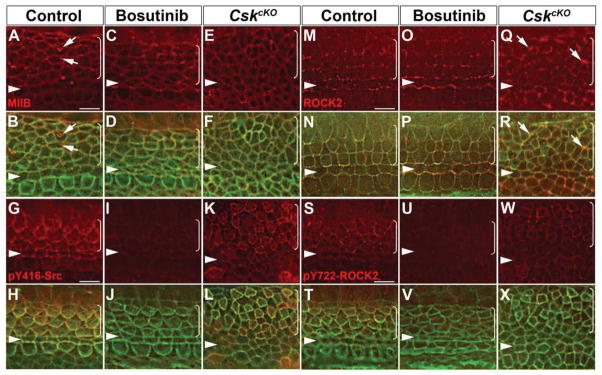
Actomyosin contractility plays a key role in tissue morphogenesis. During mammalian development, PTK7 regulates epithelial morphogenesis and planar cell polarity (PCP) through modulation of actomyosin contractility, but the underlying mechanism is unknown. Here, we show that PTK7 interacts with the tyrosine kinase Src and stimulates Src signaling along cell-cell contacts. We further identify ROCK2 as a target of junctional PTK7-Src signaling. PTK7 knockdown in cultured epithelial cells reduced the level of active Src at cell-cell contacts, resulting in delocalization of ROCK2 from cell-cell contacts and decreased junctional contractility, with a concomitant increase in actomyosin on the basal surface. Moreover, we present in vivo evidence that Src family kinase (SFK) activity is critical for PCP regulation in the auditory sensory epithelium and that PTK7-SFK signaling regulates tyrosine phosphorylation of junctional ROCK2. Together, these results delineate a PTK7-Src signaling module for spatial regulation of ROCK activity, actomyosin contractility, and epithelial PCP.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures



References
-
- Bardet PL, Guirao B, Paoletti C, Serman F, Leopold V, Bosveld F, Goya Y, Mirouse V, Graner F, Bellaiche Y. PTEN controls junction lengthening and stability during cell rearrangement in epithelial tissue. Dev Cell. 2013;25:534–546. - PubMed
-
- Bin-Nun N, Lichtig H, Malyarova A, Levy M, Elias S, Frank D. PTK7 modulates Wnt signaling activity via LRP6. Development. 2014;141:410–421. - PubMed
-
- Blankenship JT, Backovic ST, Sanny JS, Weitz O, Zallen JA. Multicellular rosette formation links planar cell polarity to tissue morphogenesis. Dev Cell. 2006;11:459–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
