

Natural forms of vitamin E: metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy
- PMID: 24704972
- PMCID: PMC4120831
- DOI: 10.1016/j.freeradbiomed.2014.03.035
Natural forms of vitamin E: metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy
Abstract
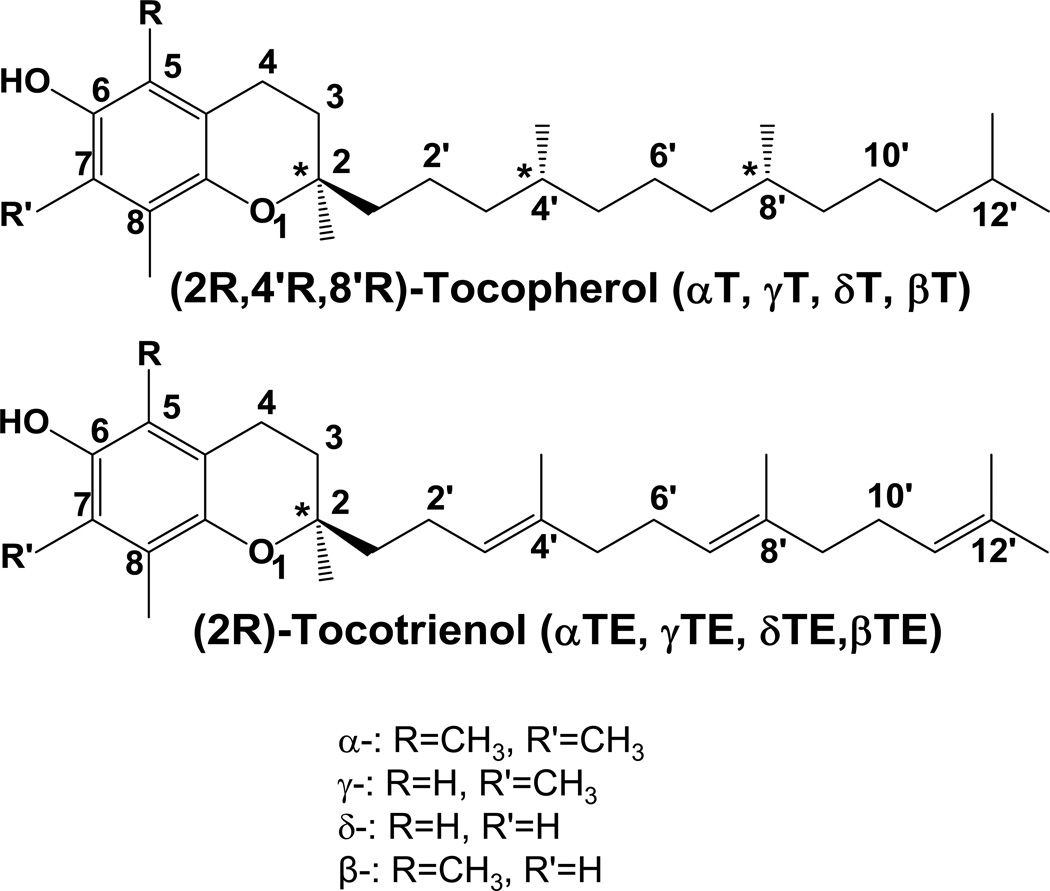
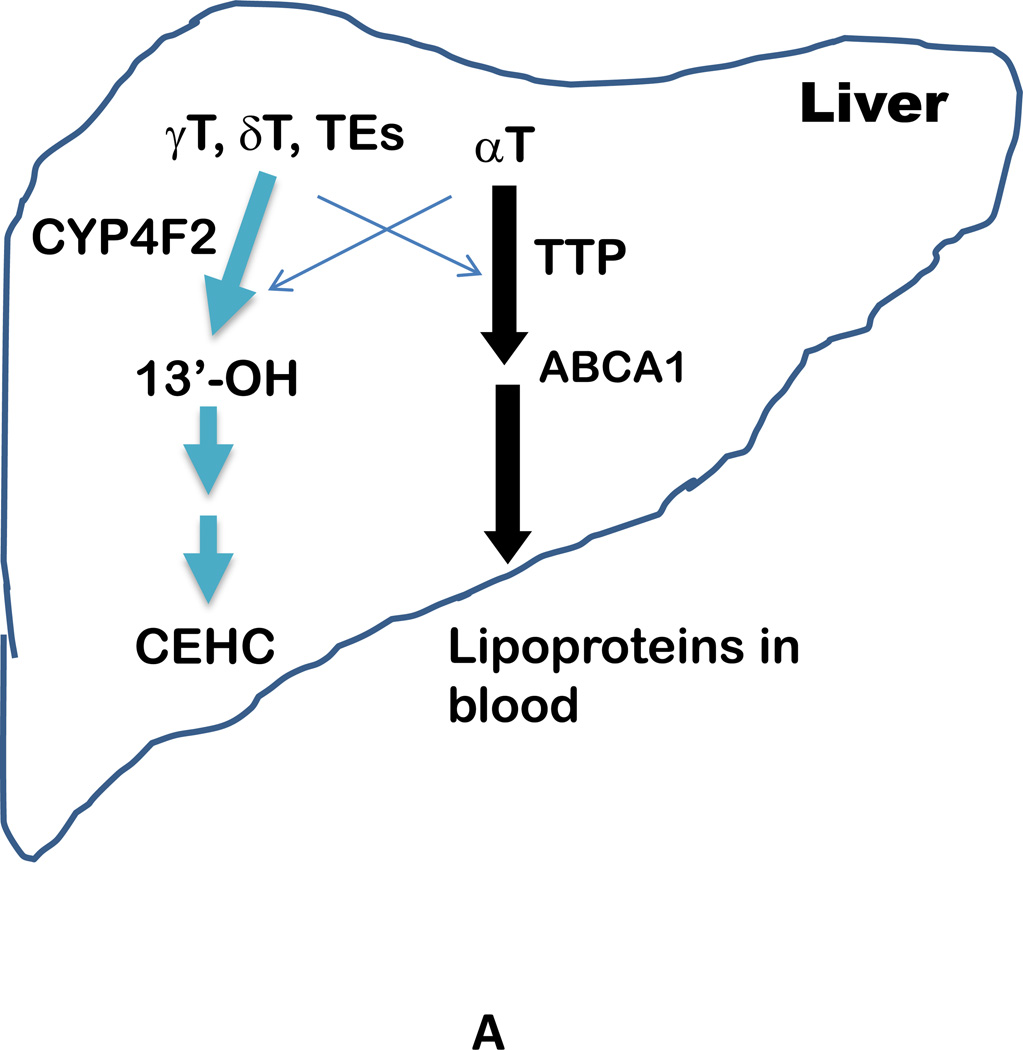
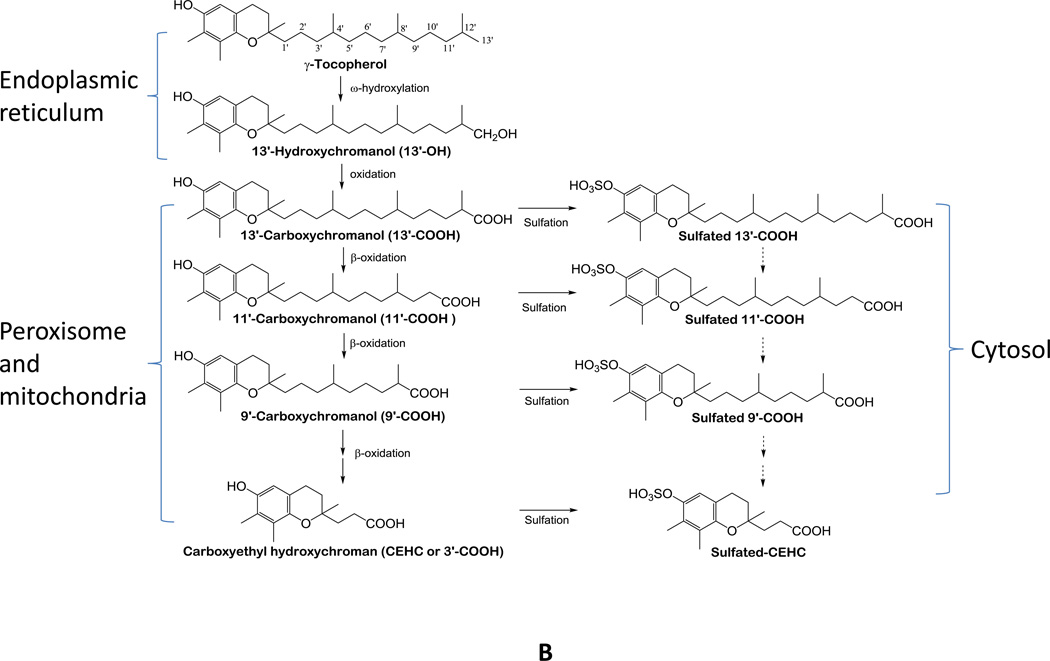
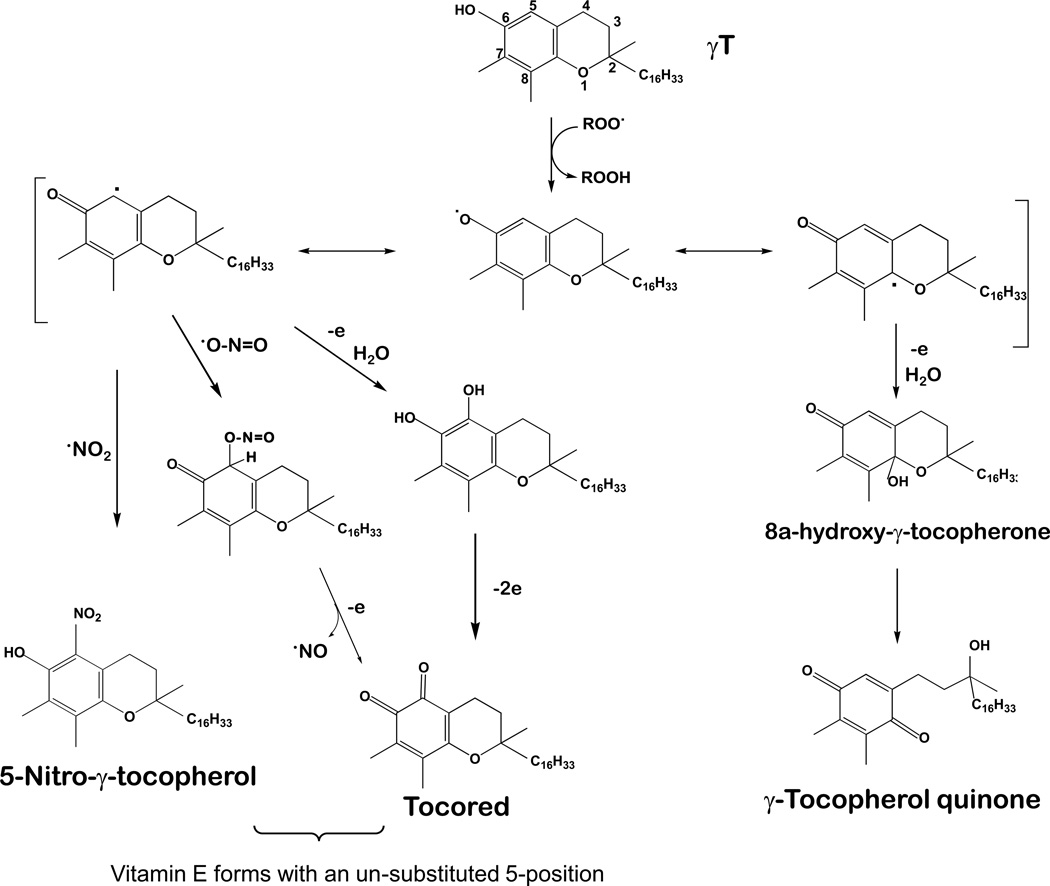
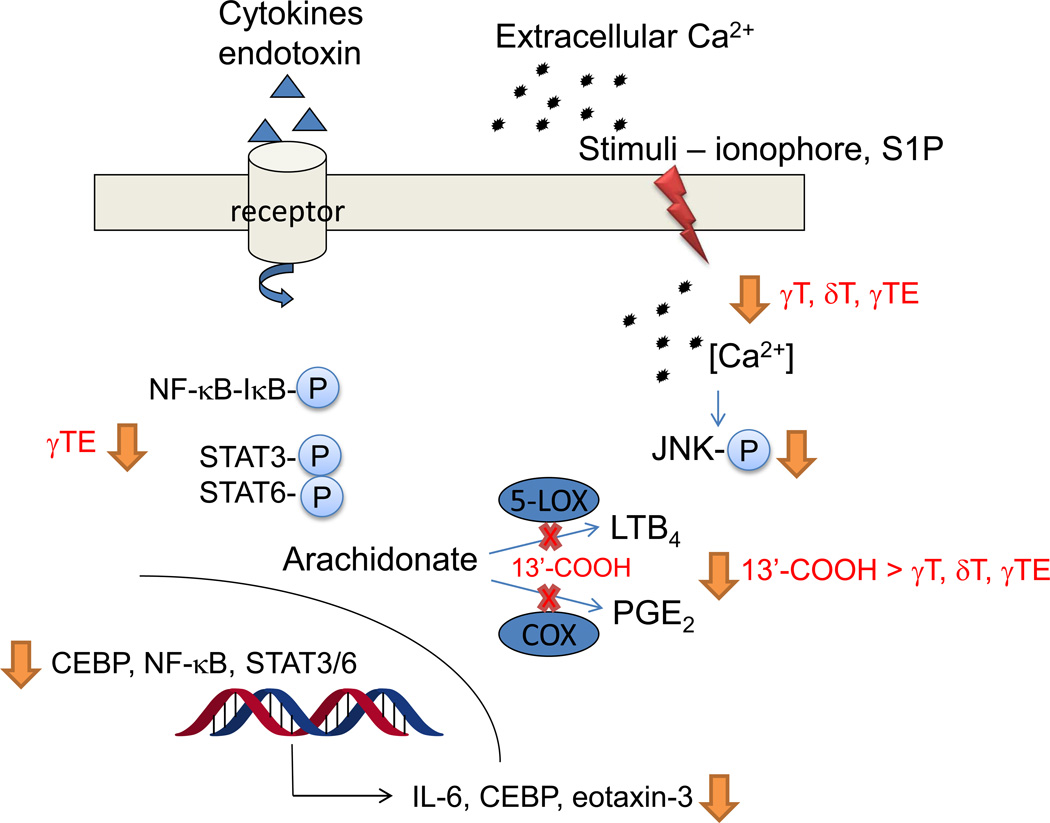
The vitamin E family consists of four tocopherols and four tocotrienols. α-Tocopherol (αT) is the predominant form of vitamin E in tissues and its deficiency leads to ataxia in humans. However, results from many clinical studies do not support a protective role of αT in disease prevention in people with adequate nutrient status. On the other hand, recent mechanistic studies indicate that other forms of vitamin E, such as γ-tocopherol (γT), δ-tocopherol, and γ-tocotrienol, have unique antioxidant and anti-inflammatory properties that are superior to those of αT in prevention and therapy against chronic diseases. These vitamin E forms scavenge reactive nitrogen species, inhibit cyclooxygenase- and 5-lipoxygenase-catalyzed eicosanoids, and suppress proinflammatory signaling such as NF-κB and STAT3/6. Unlike αT, other vitamin E forms are significantly metabolized to carboxychromanols via cytochrome P450-initiated side-chain ω-oxidation. Long-chain carboxychromanols, especially 13'-carboxychromanols, are shown to have stronger anti-inflammatory effects than unmetabolized vitamins and may therefore contribute to the beneficial effects of vitamin E forms in vivo. Consistent with mechanistic findings, animal and human studies show that γT and tocotrienols may be useful against inflammation-associated diseases. This review focuses on non-αT forms of vitamin E with respect to their metabolism, anti-inflammatory effects and mechanisms, and in vivo efficacy in preclinical models as well as human clinical intervention studies.
Keywords: 5-Lipoxygenase; Asthma; Cancer; Cyclooxygenase; Free radicals; Inflammation; Long-chain carboxychromanol; Lung injury; Tocopherol; Tocotrienol.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Jiang Q, Christen S, Shigenaga MK, Ames BN. gamma-tocopherol, the major form of vitamin E in the US diet, deserves more attention. The American journal of clinical nutrition. 2001;74:714–722. - PubMed
-
- Brigelius-Flohe R, Traber MG. Vitamin E: function and metabolism. Faseb J. 1999;13:1145–1155. - PubMed
-
- Moya-Camarena SY, Jiang Q. Sarkar Fazlul H., editor. Chapter 15-The role of vitamin E forms in cancer prevention and therapy-Studies in human intervention trials and animal models. Nutraceuticals and Cancer. 2011:323–354.
-
- Chun J, Lee J, Ye H, Exler J, Eitenmiller RR. Tocopherol and tocotrienol contents of raw and processed fruits and vegetables in the United States diet. Journal of Food Composition and Analysis. 2006;19:196–204.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
