

Design of activated serine-containing catalytic triads with atomic-level accuracy
- PMID: 24705591
- PMCID: PMC4048123
- DOI: 10.1038/nchembio.1498
Design of activated serine-containing catalytic triads with atomic-level accuracy
Abstract
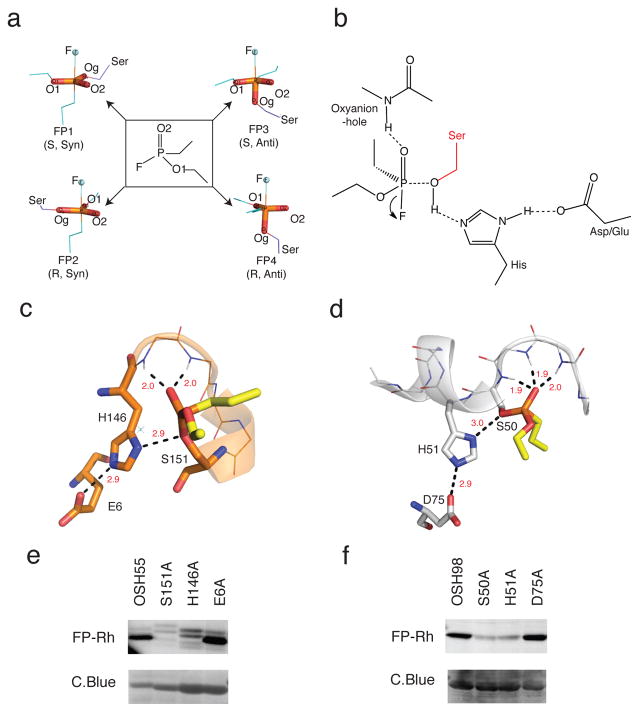
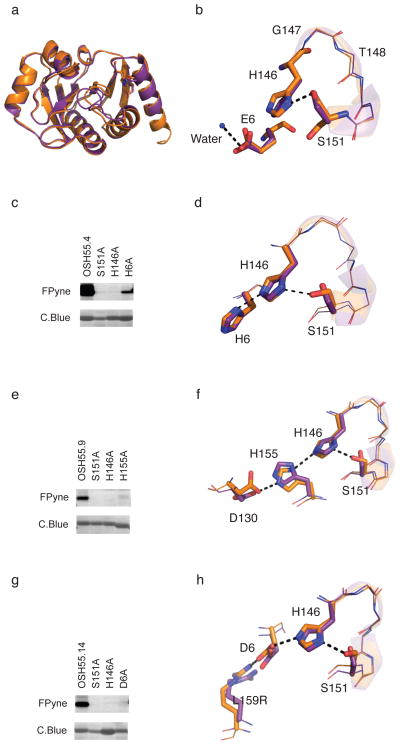
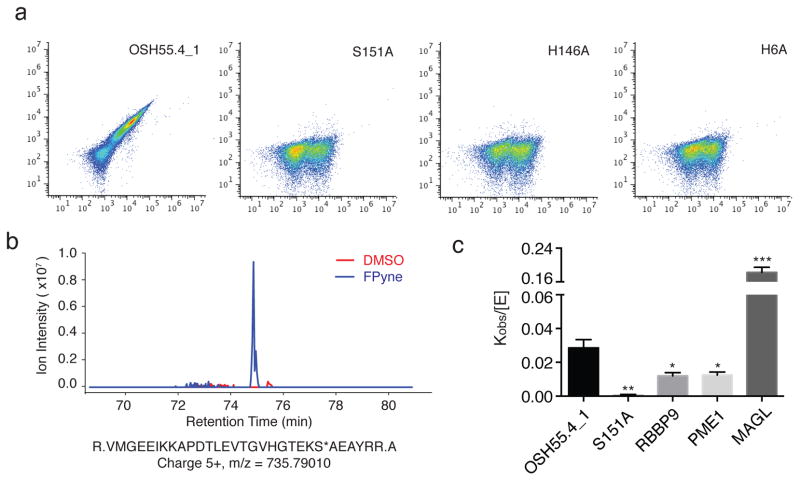
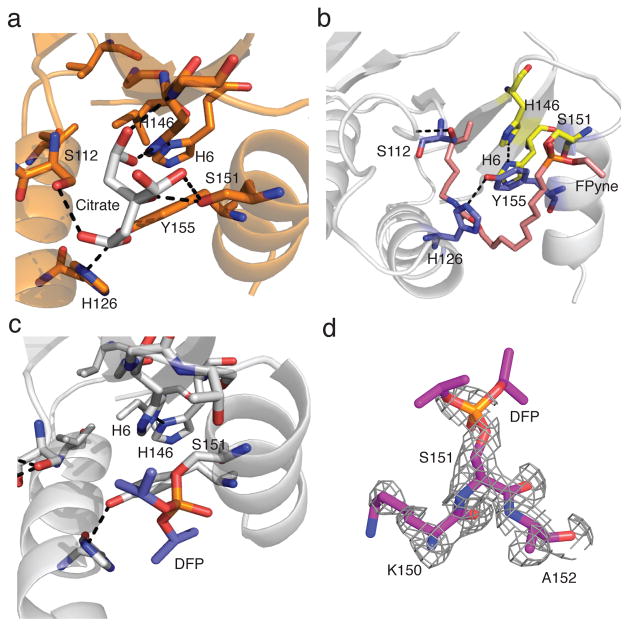
A challenge in the computational design of enzymes is that multiple properties, including substrate binding, transition state stabilization and product release, must be simultaneously optimized, and this has limited the absolute activity of successful designs. Here, we focus on a single critical property of many enzymes: the nucleophilicity of an active site residue that initiates catalysis. We design proteins with idealized serine-containing catalytic triads and assess their nucleophilicity directly in native biological systems using activity-based organophosphate probes. Crystal structures of the most successful designs show unprecedented agreement with computational models, including extensive hydrogen bonding networks between the catalytic triad (or quartet) residues, and mutagenesis experiments demonstrate that these networks are critical for serine activation and organophosphate reactivity. Following optimization by yeast display, the designs react with organophosphate probes at rates comparable to natural serine hydrolases. Co-crystal structures with diisopropyl fluorophosphate bound to the serine nucleophile suggest that the designs could provide the basis for a new class of organophosphate capture agents.
Conflict of interest statement
Figures
References
-
- Hedstrom L. Serine protease mechanism and specificity. Chem Rev. 2002;102:4501–4524. - PubMed
-
- Carter P, Wells JA. Dissecting the catalytic triad of a serine protease. Nature. 1988;332:564–568. - PubMed
-
- Corey DR, Craik CS. An Investigation into the Minimum Requirements for Peptide Hydrolysis by Mutation of the Catalytic Traid of Trypsin. J Am Chem Soc. 1992;114:1784–1790.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
