

Integrated strategy reveals the protein interface between cancer targets Bcl-2 and NAF-1
- PMID: 24706857
- PMCID: PMC3986192
- DOI: 10.1073/pnas.1403770111
Integrated strategy reveals the protein interface between cancer targets Bcl-2 and NAF-1
Abstract
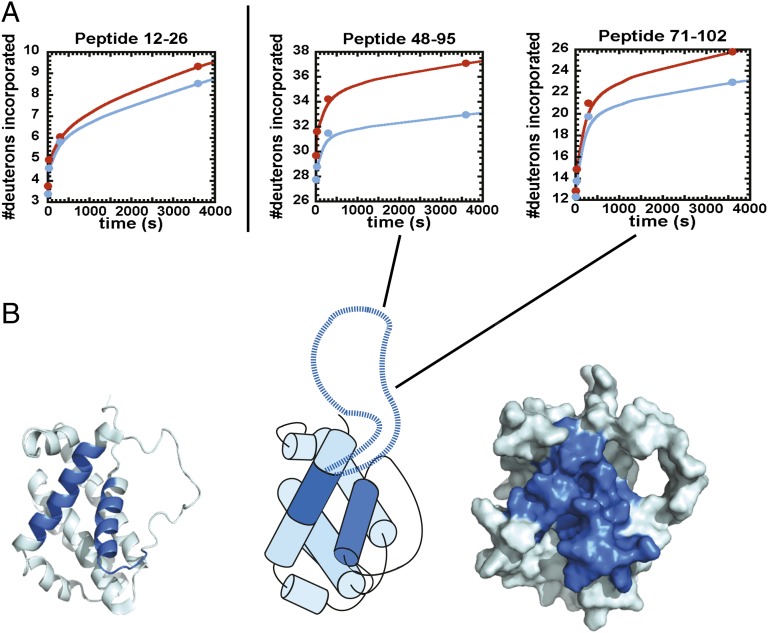
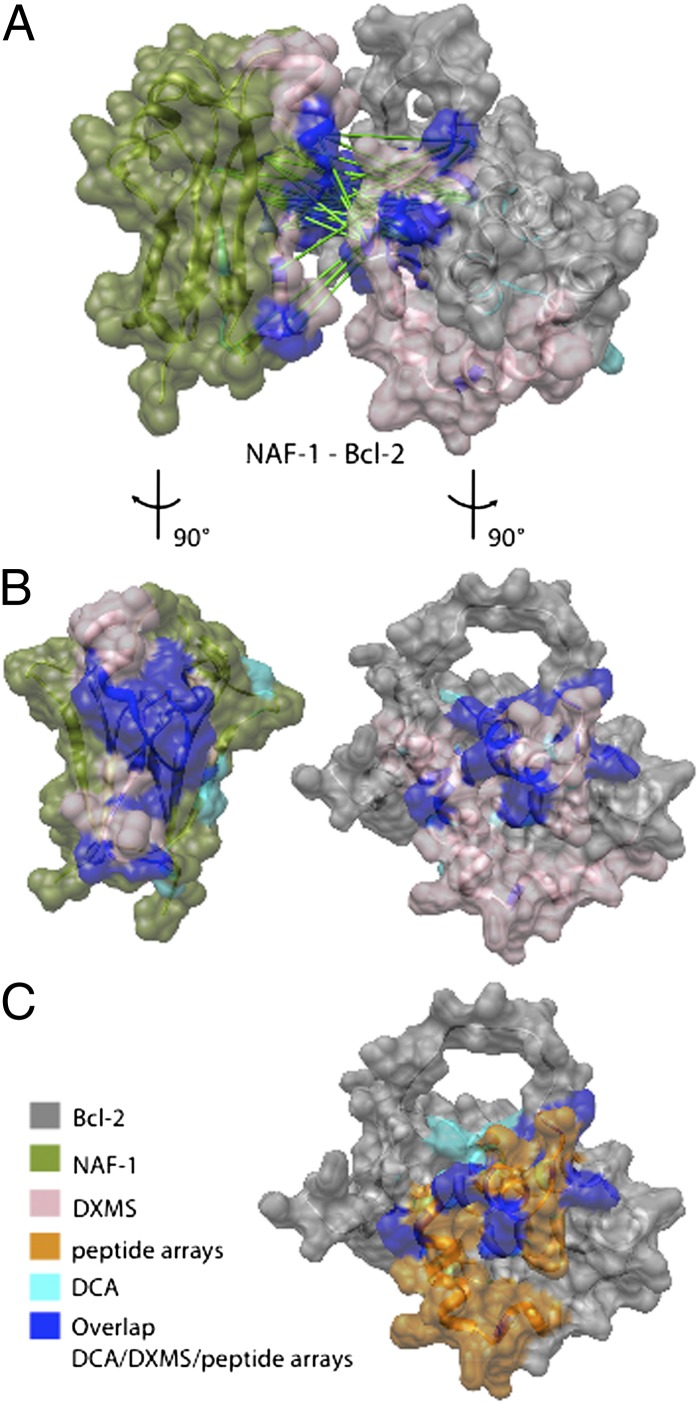
Life requires orchestrated control of cell proliferation, cell maintenance, and cell death. Involved in these decisions are protein complexes that assimilate a variety of inputs that report on the status of the cell and lead to an output response. Among the proteins involved in this response are nutrient-deprivation autophagy factor-1 (NAF-1)- and Bcl-2. NAF-1 is a homodimeric member of the novel Fe-S protein NEET family, which binds two 2Fe-2S clusters. NAF-1 is an important partner for Bcl-2 at the endoplasmic reticulum to functionally antagonize Beclin 1-dependent autophagy [Chang NC, Nguyen M, Germain M, Shore GC (2010) EMBO J 29(3):606-618]. We used an integrated approach involving peptide array, deuterium exchange mass spectrometry (DXMS), and functional studies aided by the power of sufficient constraints from direct coupling analysis (DCA) to determine the dominant docked conformation of the NAF-1-Bcl-2 complex. NAF-1 binds to both the pro- and antiapoptotic regions (BH3 and BH4) of Bcl-2, as demonstrated by a nested protein fragment analysis in a peptide array and DXMS analysis. A combination of the solution studies together with a new application of DCA to the eukaryotic proteins NAF-1 and Bcl-2 provided sufficient constraints at amino acid resolution to predict the interaction surfaces and orientation of the protein-protein interactions involved in the docked structure. The specific integrated approach described in this paper provides the first structural information, to our knowledge, for future targeting of the NAF-1-Bcl-2 complex in the regulation of apoptosis/autophagy in cancer biology.
Keywords: CDGSH; Cisd1; Cisd2; Miner1; mitoNEET.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
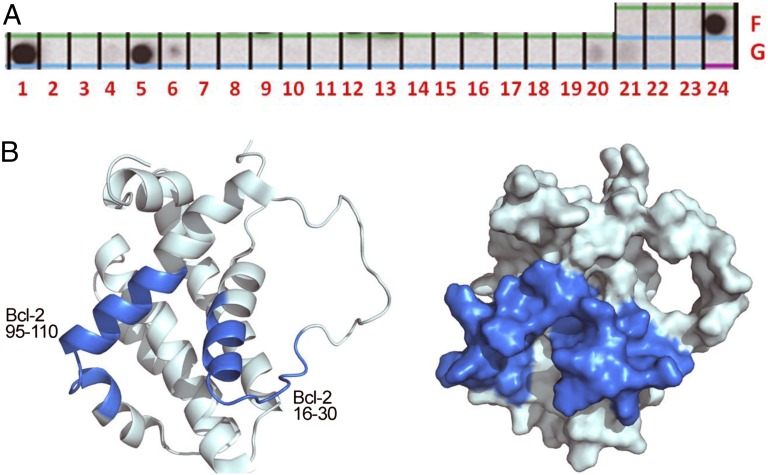
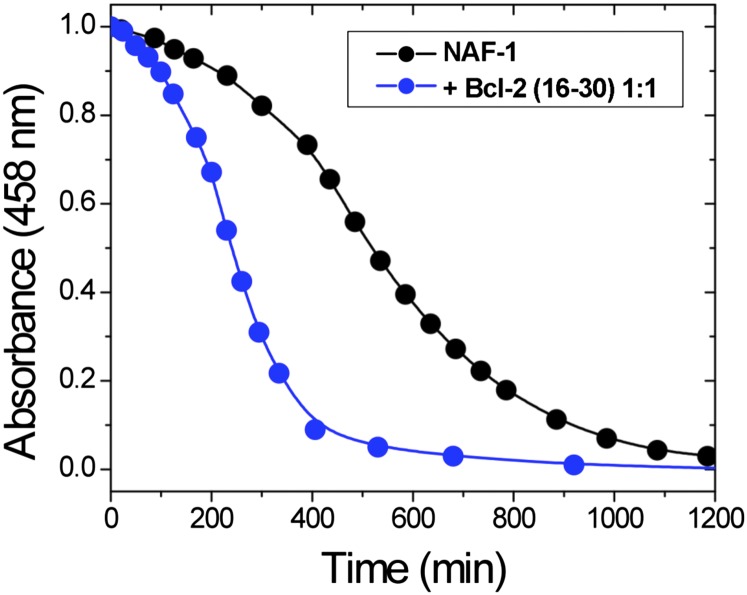
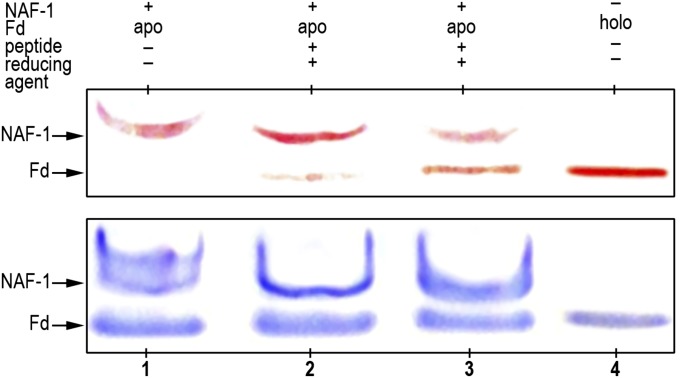

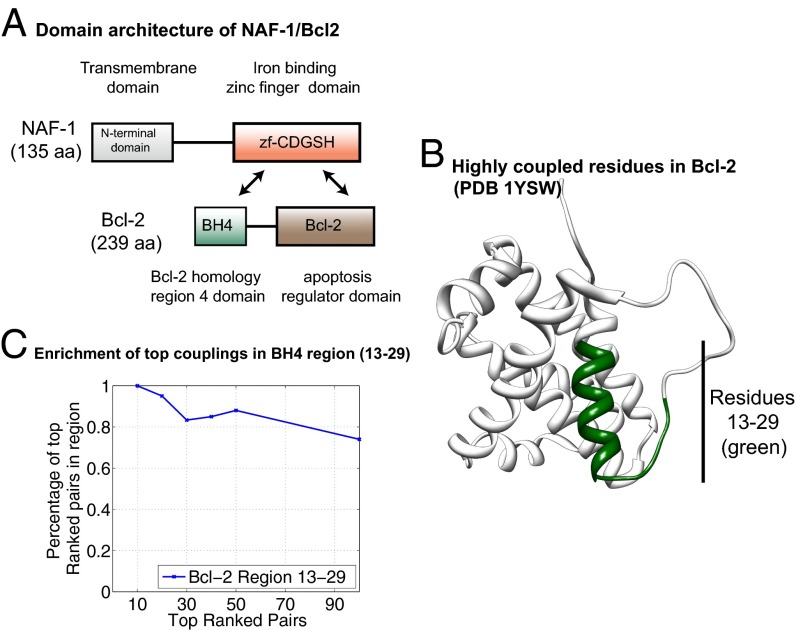
References
-
- Pattingre S, et al. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell. 2005;122(6):927–939. - PubMed
-
- Youle RJ, Strasser A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat Rev Mol Cell Biol. 2008;9(1):47–59. - PubMed
-
- Chang NC, et al. Bcl-2-associated autophagy regulator Naf-1 required for maintenance of skeletal muscle. Hum Mol Genet. 2012;21(10):2277–2287. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
