

GAS2-like proteins mediate communication between microtubules and actin through interactions with end-binding proteins
- PMID: 24706950
- PMCID: PMC4058111
- DOI: 10.1242/jcs.140558
GAS2-like proteins mediate communication between microtubules and actin through interactions with end-binding proteins
Abstract
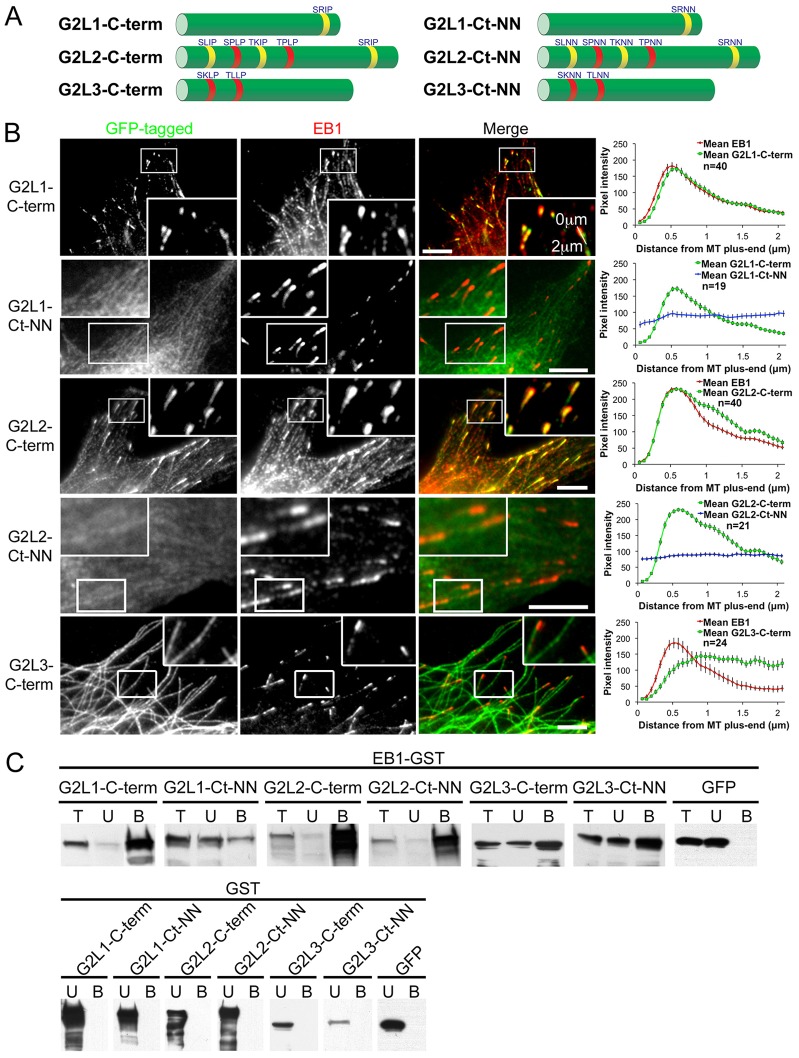
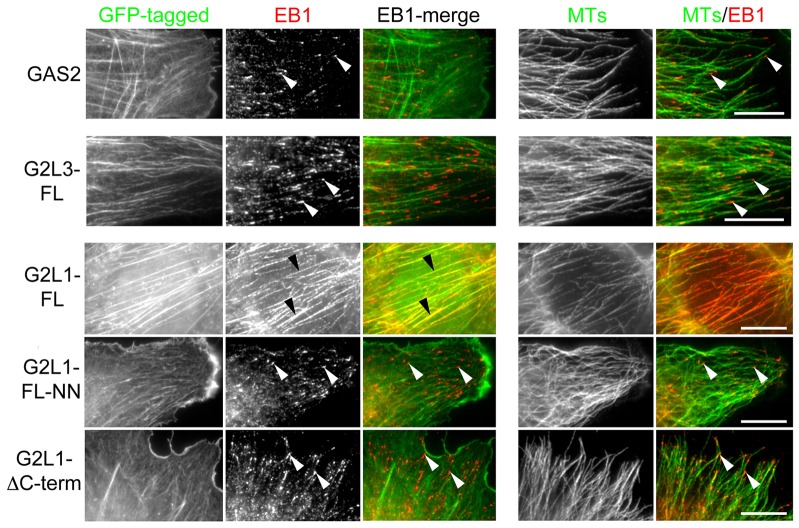
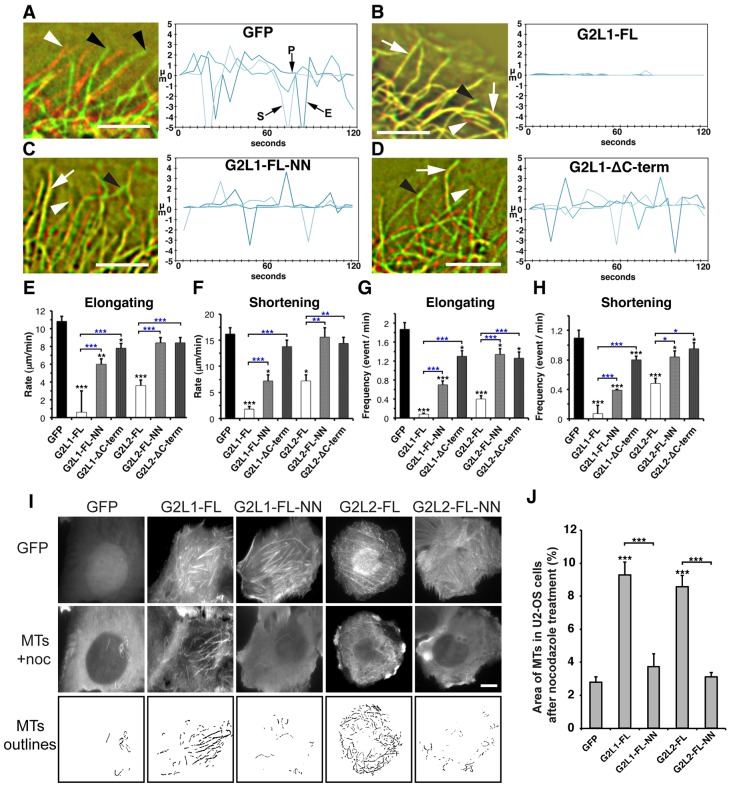
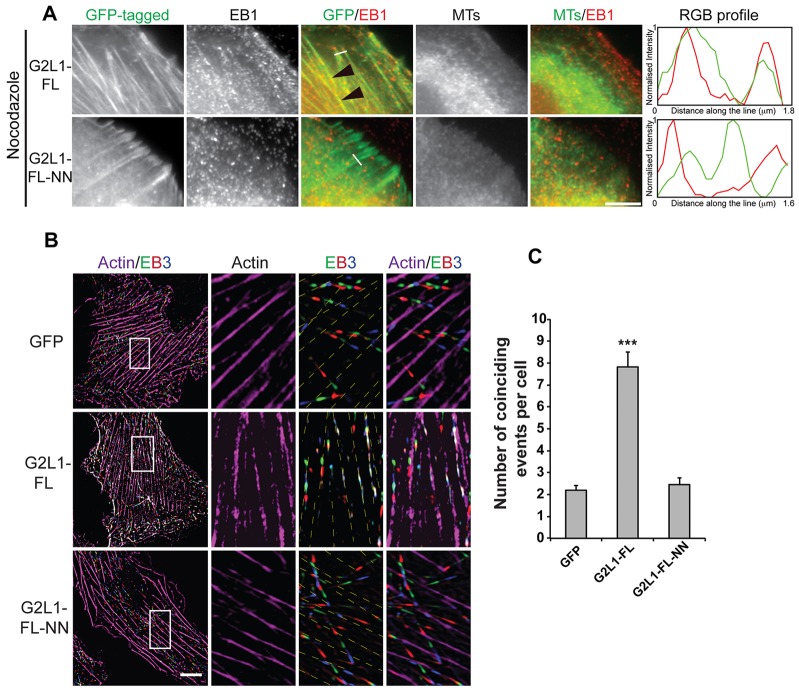
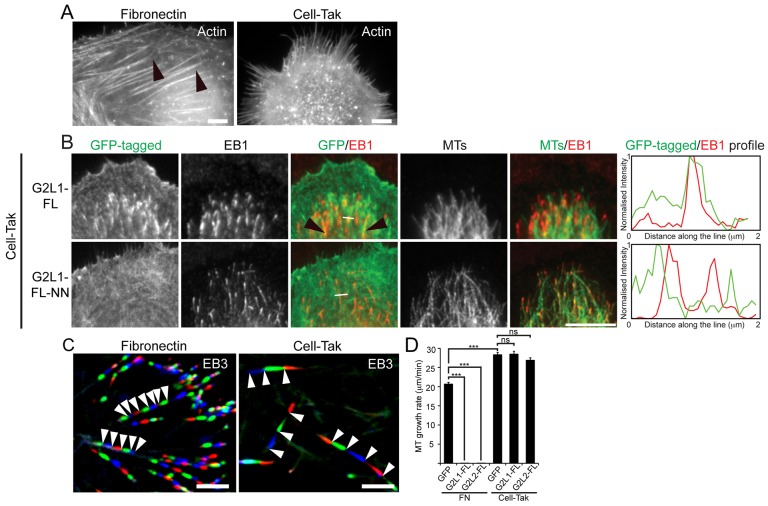
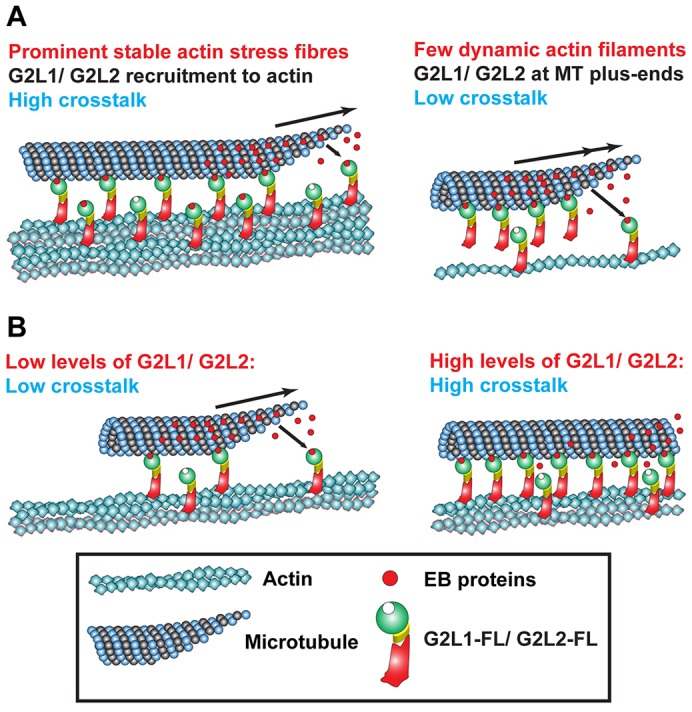
Crosstalk between the microtubule (MT) and actin cytoskeletons is fundamental to many cellular processes including cell polarisation and cell motility. Previous work has shown that members of the growth-arrest-specific 2 (GAS2) family mediate the crosstalk between filamentous actin (F-actin) and MTs, but the molecular basis of this process remained unclear. By using fluorescence microscopy, we demonstrate that three members of this family, GAS2-like 1, GAS2-like 2 and GAS2-like 3 (G2L1, G2L2 and G2L3, also known as GAS2L1, GAS2L2 and GAS2L3, respectively) are differentially involved in mediating the crosstalk between F-actin and MTs. Although all localise to actin and MTs, only the exogenous expression of G2L1 and G2L2 influenced MT stability, dynamics and guidance along actin stress fibres. Biochemical analysis and live-cell imaging revealed that their functions are largely due to the association of these proteins with MT plus-end-binding proteins that bind to SxIP or SxLP motifs located at G2L C-termini. Our findings lead to a model in which end-binding (EB) proteins play a key role in mediating actin-MT crosstalk.
Keywords: Actin; End-binding protein; GAS2 family; GAS2-like 1; GAS2-like 2; GAS2-like 3; MT-tip localising signal; Microtubule; MtLS.
© 2014. Published by The Company of Biologists Ltd.
Figures

References
-
- Alves-Silva J., Sanchez-Soriano N., Beaven R., Klein M., Parkin J., Millard T. H., Bellen H. J., Venken K. J., Ballestrem C., Kammerer R. A. et al. (2012). Spectraplakins promote microtubule-mediated axonal growth by functioning as structural microtubule-associated proteins and EB1-dependent +TIPs (tip interacting proteins). J. Neurosci. 32, 9143–9158 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
