

cAMP-PKA phosphorylation of tau confers risk for degeneration in aging association cortex
- PMID: 24707050
- PMCID: PMC3977284
- DOI: 10.1073/pnas.1322360111
cAMP-PKA phosphorylation of tau confers risk for degeneration in aging association cortex
Abstract
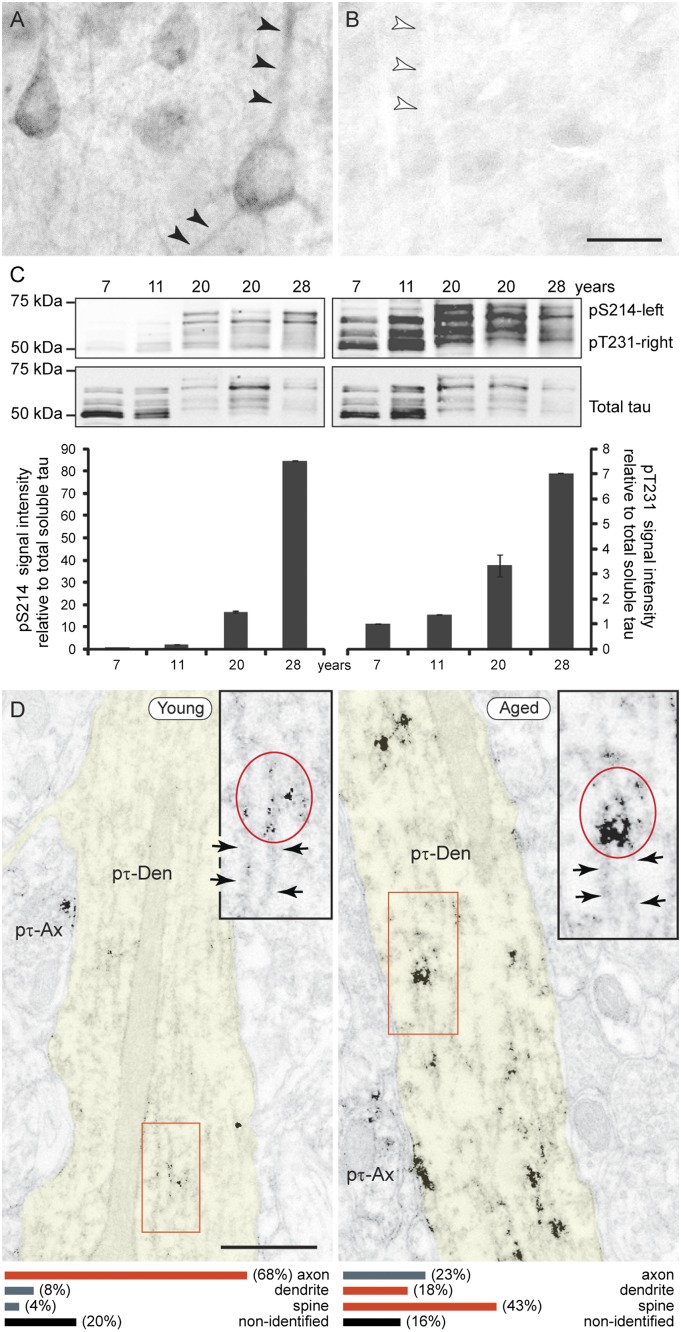
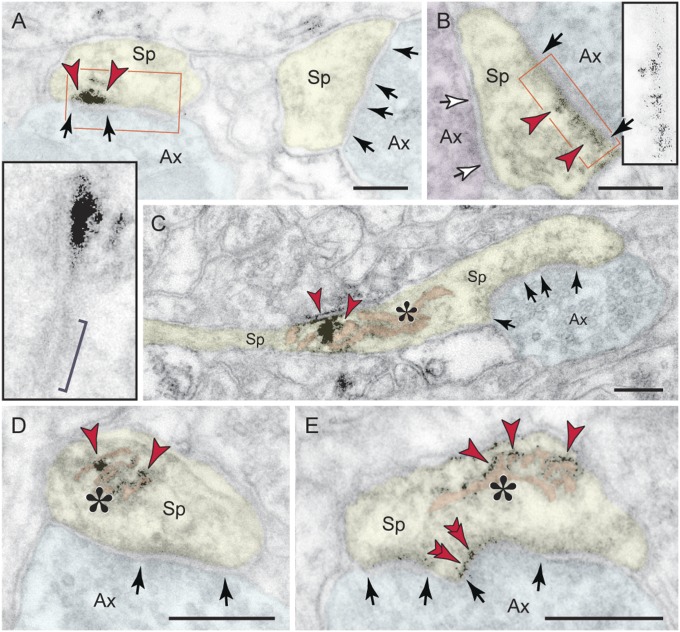
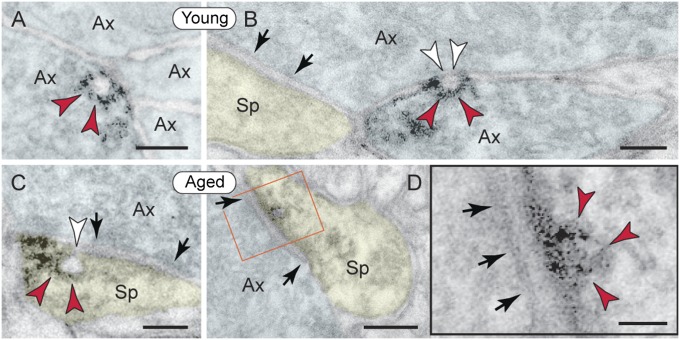
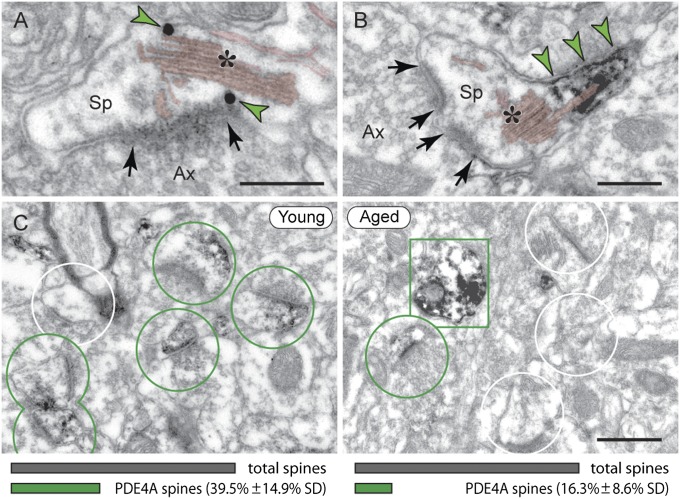
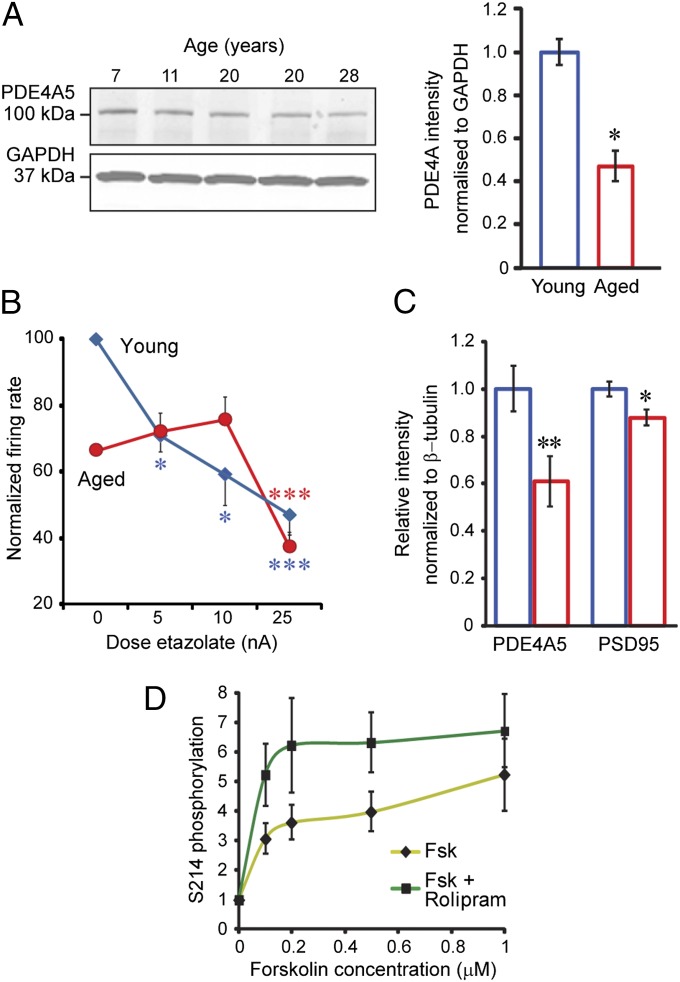
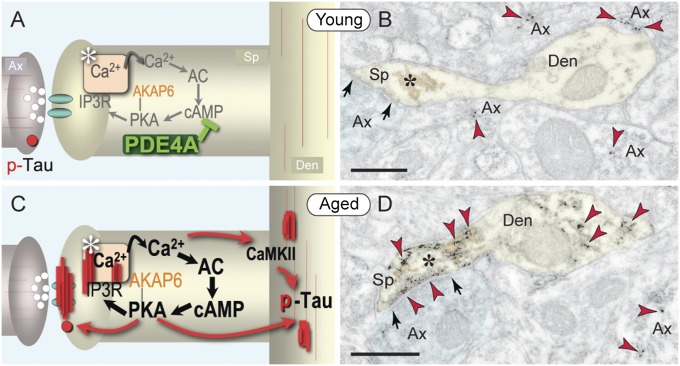
The pattern of neurodegeneration in Alzheimer's disease (AD) is very distinctive: neurofibrillary tangles (NFTs) composed of hyperphosphorylated tau selectively affect pyramidal neurons of the aging association cortex that interconnect extensively through glutamate synapses on dendritic spines. In contrast, primary sensory cortices have few NFTs, even in late-stage disease. Understanding this selective vulnerability, and why advancing age is such a high risk factor for the degenerative process, may help to reveal disease etiology and provide targets for intervention. Our study has revealed age-related increase in cAMP-dependent protein kinase (PKA) phosphorylation of tau at serine 214 (pS214-tau) in monkey dorsolateral prefrontal association cortex (dlPFC), which specifically targets spine synapses and the Ca(2+)-storing spine apparatus. This increase is mirrored by loss of phosphodiesterase 4A from the spine apparatus, consistent with increase in cAMP-Ca(2+) signaling in aging spines. Phosphorylated tau was not detected in primary visual cortex, similar to the pattern observed in AD. We also report electron microscopic evidence of previously unidentified vesicular trafficking of phosphorylated tau in normal association cortex--in axons in young dlPFC vs. in spines in aged dlPFC--consistent with the transneuronal lesion spread reported in genetic rodent models. pS214-Tau was not observed in normal aged mice, suggesting that it arises with the evolutionary expansion of corticocortical connections in primates, crossing the threshold into NFTs and degeneration in humans. Thus, the cAMP-Ca(2+) signaling mechanisms, needed for flexibly modulating network strength in young association cortex, confer vulnerability to degeneration when dysregulated with advancing age.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Interrogating the Etiology of Sporadic Alzheimer's Disease Using Aging Rhesus Macaques: Cellular, Molecular, and Cortical Circuitry Perspectives.J Gerontol A Biol Sci Med Sci. 2023 Aug 27;78(9):1523-1534. doi: 10.1093/gerona/glad134. J Gerontol A Biol Sci Med Sci. 2023. PMID: 37279946 Free PMC article.
-
Age-related calcium dysregulation linked with tau pathology and impaired cognition in non-human primates.Alzheimers Dement. 2021 Jun;17(6):920-932. doi: 10.1002/alz.12325. Epub 2021 Apr 7. Alzheimers Dement. 2021. PMID: 33829643 Free PMC article.
-
Synaptic distributions of pS214-tau in rhesus monkey prefrontal cortex are associated with spine density, but not with cognitive decline.J Comp Neurol. 2019 Mar 1;527(4):856-873. doi: 10.1002/cne.24576. Epub 2018 Dec 4. J Comp Neurol. 2019. PMID: 30408169 Free PMC article.
-
Studies of aging nonhuman primates illuminate the etiology of early-stage Alzheimer's-like neuropathology: An evolutionary perspective.Am J Primatol. 2021 Nov;83(11):e23254. doi: 10.1002/ajp.23254. Epub 2021 May 7. Am J Primatol. 2021. PMID: 33960505 Free PMC article. Review.
-
[Drug development for tauopathy and Alzheimer's disease].Nihon Shinkei Seishin Yakurigaku Zasshi. 2010 Aug;30(4):177-80. Nihon Shinkei Seishin Yakurigaku Zasshi. 2010. PMID: 20857696 Review. Japanese.
Cited by
-
The Oral-Gut-Brain AXIS: The Influence of Microbes in Alzheimer's Disease.Front Cell Neurosci. 2021 Apr 14;15:633735. doi: 10.3389/fncel.2021.633735. eCollection 2021. Front Cell Neurosci. 2021. PMID: 33935651 Free PMC article. Review.
-
Simple, Single-Shot Phosphoproteomic Analysis of Heat-Stable Tau Identifies Age-Related Changes in pS235- and pS396-Tau Levels in Non-human Primates.Front Aging Neurosci. 2021 Nov 18;13:767322. doi: 10.3389/fnagi.2021.767322. eCollection 2021. Front Aging Neurosci. 2021. PMID: 34867294 Free PMC article.
-
Interrogating the Etiology of Sporadic Alzheimer's Disease Using Aging Rhesus Macaques: Cellular, Molecular, and Cortical Circuitry Perspectives.J Gerontol A Biol Sci Med Sci. 2023 Aug 27;78(9):1523-1534. doi: 10.1093/gerona/glad134. J Gerontol A Biol Sci Med Sci. 2023. PMID: 37279946 Free PMC article.
-
Bacillus bombysepticus α-Toxin Binding to G Protein-Coupled Receptor Kinase 2 Regulates cAMP/PKA Signaling Pathway to Induce Host Death.PLoS Pathog. 2016 Mar 29;12(3):e1005527. doi: 10.1371/journal.ppat.1005527. eCollection 2016 Mar. PLoS Pathog. 2016. PMID: 27022742 Free PMC article.
-
Phosphodiesterase PDE4D Is Decreased in Frontal Cortex of Aged Rats and Positively Correlated With Working Memory Performance and Inversely Correlated With PKA Phosphorylation of Tau.Front Aging Neurosci. 2020 Oct 28;12:576723. doi: 10.3389/fnagi.2020.576723. eCollection 2020. Front Aging Neurosci. 2020. PMID: 33192469 Free PMC article.
References
-
- Evans DA, et al. Prevalence of Alzheimer’s disease in a community population of older persons. Higher than previously reported. JAMA. 1989;262(18):2551–2556. - PubMed
-
- Braak H, Thal DR, Ghebremedhin E, Del Tredici K. Stages of the pathologic process in Alzheimer disease: Age categories from 1 to 100 years. J Neuropathol Exp Neurol. 2011;70(11):960–969. - PubMed
-
- Giannakopoulos P, et al. Tangle and neuron numbers, but not amyloid load, predict cognitive status in Alzheimer’s disease. Neurology. 2003;60(9):1495–1500. - PubMed
-
- Sengupta A, et al. Phosphorylation of tau at both Thr 231 and Ser 262 is required for maximal inhibition of its binding to microtubules. Arch Biochem Biophys. 1998;357(2):299–309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
