

PDF and cAMP enhance PER stability in Drosophila clock neurons
- PMID: 24707054
- PMCID: PMC3977231
- DOI: 10.1073/pnas.1402562111
PDF and cAMP enhance PER stability in Drosophila clock neurons
Erratum in
- Proc Natl Acad Sci U S A. 2014 Jun 3;111(22):8311
Abstract
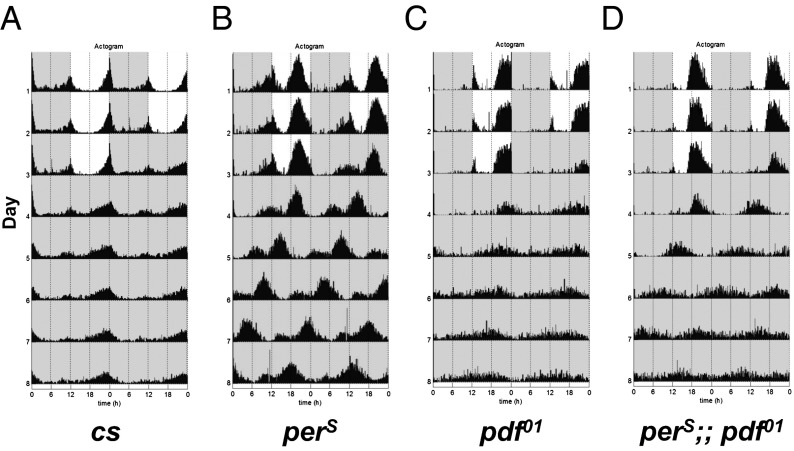
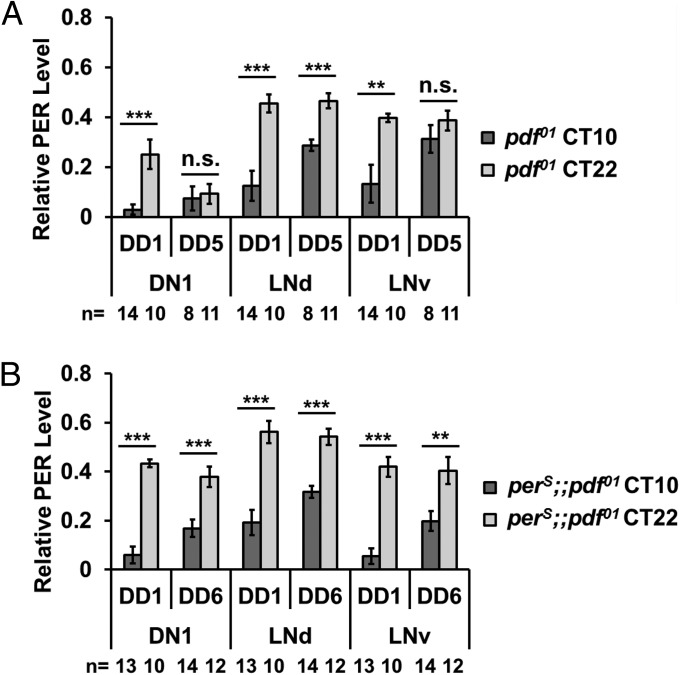
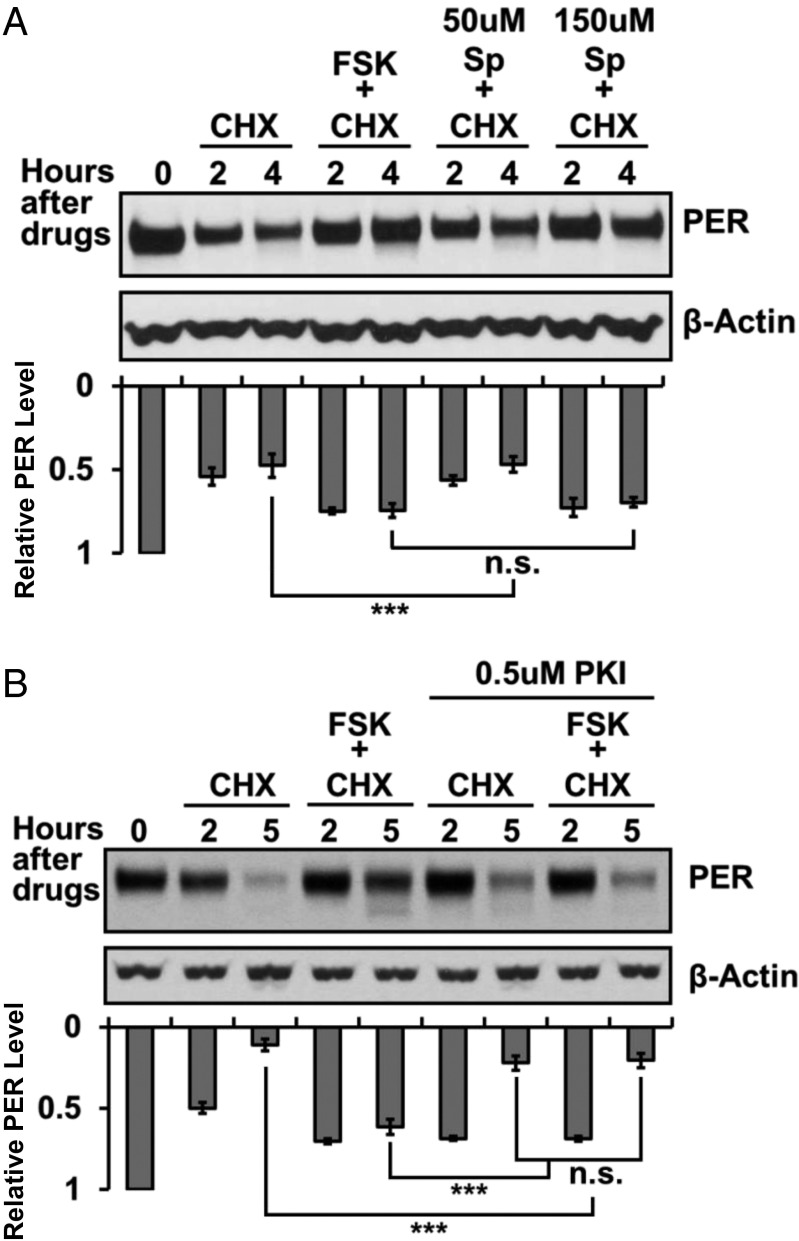
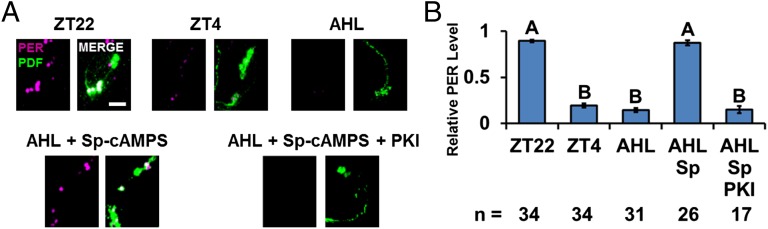
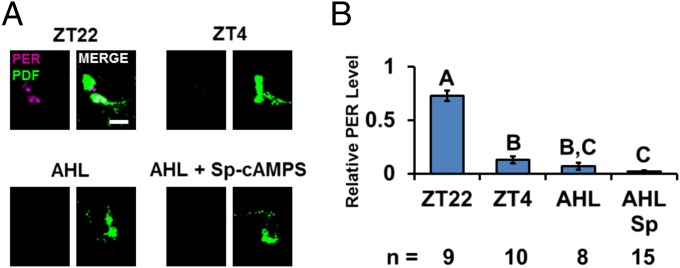
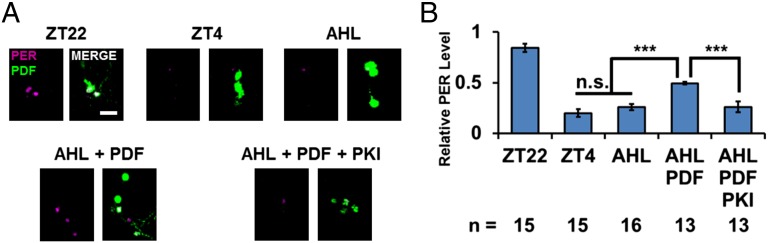
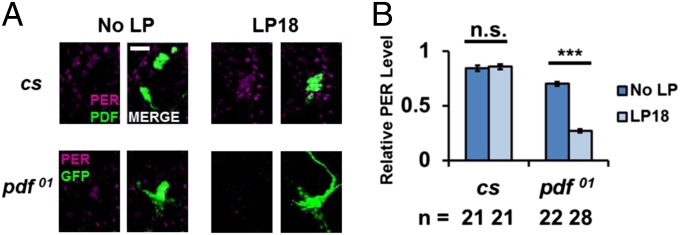
The neuropeptide PDF is important for Drosophila circadian rhythms: pdf(01) (pdf-null) animals are mostly arrhythmic or short period in constant darkness and have an advanced activity peak in light-dark conditions. PDF contributes to the amplitude, synchrony, as well as the pace of circadian rhythms within clock neurons. PDF is known to increase cAMP levels in PDR receptor (PDFR)-containing neurons. However, there is no known connection of PDF or of cAMP with the Drosophila molecular clockworks. We discovered that the mutant period gene per(S) ameliorates the phenotypes of pdf-null flies. The period protein (PER) is a well-studied repressor of clock gene transcription, and the per(S) protein (PERS) has a markedly short half-life. The result therefore suggests that the PDF-mediated increase in cAMP might lengthen circadian period by directly enhancing PER stability. Indeed, increasing cAMP levels and cAMP-mediated protein kinase A (PKA) activity stabilizes PER, in S2 tissue culture cells and in fly circadian neurons. Adding PDF to fly brains in vitro has a similar effect. Consistent with these relationships, a light pulse causes more prominent PER degradation in pdf(01) circadian neurons than in wild-type neurons. The results indicate that PDF contributes to clock neuron synchrony by increasing cAMP and PKA, which enhance PER stability and decrease clock speed in intrinsically fast-paced PDFR-containing clock neurons. We further suggest that the more rapid degradation of PERS bypasses PKA regulation and makes the pace of clock neurons more uniform, allowing them to avoid much of the asynchrony caused by the absence of PDF.
Keywords: PDF neurons; PDF signaling; molecular clock regulation; synchronization.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Circadian Rhythms and Sleep in Drosophila melanogaster.Genetics. 2017 Apr;205(4):1373-1397. doi: 10.1534/genetics.115.185157. Genetics. 2017. PMID: 28360128 Free PMC article. Review.
-
The Drosophila Receptor Protein Tyrosine Phosphatase LAR Is Required for Development of Circadian Pacemaker Neuron Processes That Support Rhythmic Activity in Constant Darkness But Not during Light/Dark Cycles.J Neurosci. 2016 Mar 30;36(13):3860-70. doi: 10.1523/JNEUROSCI.4523-15.2016. J Neurosci. 2016. PMID: 27030770 Free PMC article.
-
Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain.Nature. 2004 Oct 14;431(7010):869-73. doi: 10.1038/nature02935. Nature. 2004. PMID: 15483616
-
Electrical silencing of PDF neurons advances the phase of non-PDF clock neurons in Drosophila.J Biol Rhythms. 2008 Apr;23(2):117-28. doi: 10.1177/0748730407312984. J Biol Rhythms. 2008. PMID: 18375861
-
Circadian clock genes in Drosophila: recent developments.Indian J Exp Biol. 2003 Aug;41(8):797-804. Indian J Exp Biol. 2003. PMID: 15248475 Review.
Cited by
-
Codon usage affects the structure and function of the Drosophila circadian clock protein PERIOD.Genes Dev. 2016 Aug 1;30(15):1761-75. doi: 10.1101/gad.281030.116. Genes Dev. 2016. PMID: 27542830 Free PMC article.
-
Functional PDF Signaling in the Drosophila Circadian Neural Circuit Is Gated by Ral A-Dependent Modulation.Neuron. 2016 May 18;90(4):781-794. doi: 10.1016/j.neuron.2016.04.002. Epub 2016 May 5. Neuron. 2016. PMID: 27161526 Free PMC article.
-
Pigment-Dispersing Factor Signaling and Circadian Rhythms in Insect Locomotor Activity.Curr Opin Insect Sci. 2014 Jul 1;1:73-80. doi: 10.1016/j.cois.2014.05.002. Curr Opin Insect Sci. 2014. PMID: 25386391 Free PMC article.
-
Circadian Rhythms and Sleep in Drosophila melanogaster.Genetics. 2017 Apr;205(4):1373-1397. doi: 10.1534/genetics.115.185157. Genetics. 2017. PMID: 28360128 Free PMC article. Review.
-
MicroRNA-92a is a circadian modulator of neuronal excitability in Drosophila.Nat Commun. 2017 Mar 9;8:14707. doi: 10.1038/ncomms14707. Nat Commun. 2017. PMID: 28276426 Free PMC article.
References
-
- Hardin PE, Hall JC, Rosbash M. Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature. 1990;343(6258):536–540. - PubMed
-
- Sehgal A, et al. Rhythmic expression of timeless: A basis for promoting circadian cycles in period gene autoregulation. Science. 1995;270(5237):808–810. - PubMed
-
- Darlington TK, et al. Closing the circadian loop: CLOCK-induced transcription of its own inhibitors per and tim. Science. 1998;280(5369):1599–1603. - PubMed
-
- Renn SC, Park JH, Rosbash M, Hall JC, Taghert PH. A pdf neuropeptide gene mutation and ablation of PDF neurons each cause severe abnormalities of behavioral circadian rhythms in Drosophila. Cell. 1999;99(7):791–802. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
