

Somatic hypermutation at A/T-rich oligonucleotide substrates shows different strand polarities in Ung-deficient or -proficient backgrounds
- PMID: 24710273
- PMCID: PMC4054293
- DOI: 10.1128/MCB.01452-13
Somatic hypermutation at A/T-rich oligonucleotide substrates shows different strand polarities in Ung-deficient or -proficient backgrounds
Abstract
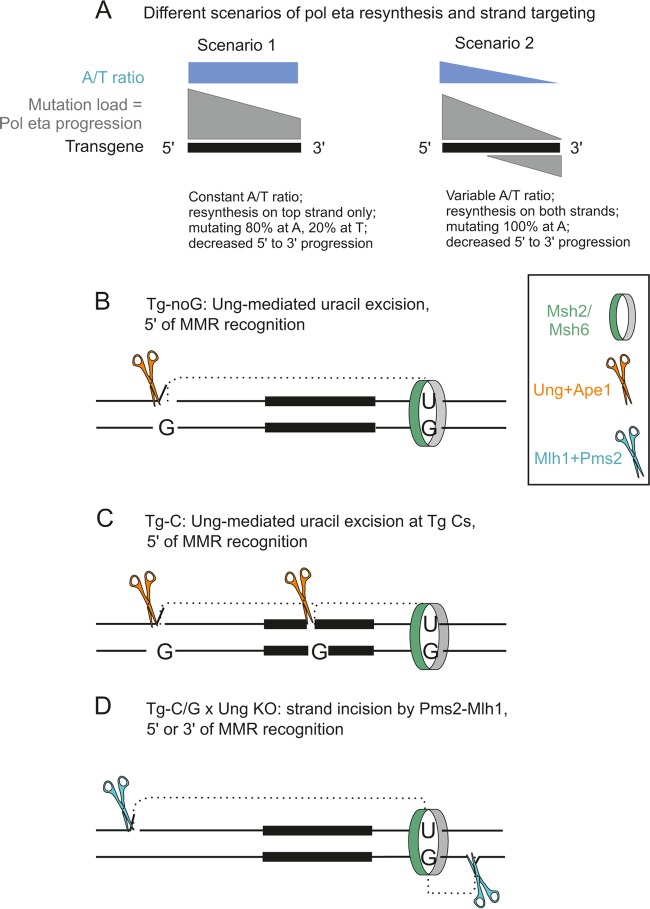
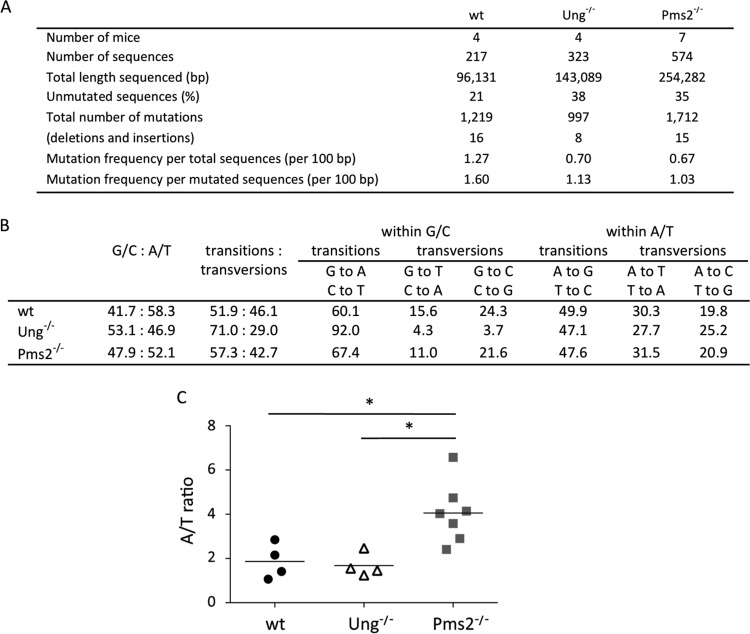
A/T mutations at immunoglobulin loci are introduced by DNA polymerase η (Polη) during an Msh2/6-dependent repair process which results in A's being mutated 2-fold more often than T's. This patch synthesis is initiated by a DNA incision event whose origin is still obscure. We report here the analysis of A/T oligonucleotide mutation substrates inserted at the heavy chain locus, including or not including internal C's or G's. Surprisingly, the template composed of only A's and T's was highly mutated over its entire 90-bp length, with a 2-fold decrease in mutation from the 5' to the 3' end and a constant A/T ratio of 4. These results imply that Polη synthesis was initiated from a break in the 5'-flanking region of the substrate and proceeded over its entire length. The A/T bias was strikingly altered in an Ung(-/-) background, which provides the first experimental evidence supporting a concerted action of Ung and Msh2/6 pathways to generate mutations at A/T bases. New analysis of Pms2(-/-) animals provided a complementary picture, revealing an A/T mutation ratio of 4. We therefore propose that Ung and Pms2 may exert a mutual backup function for the DNA incision that promotes synthesis by Polη, each with a distinct strand bias.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
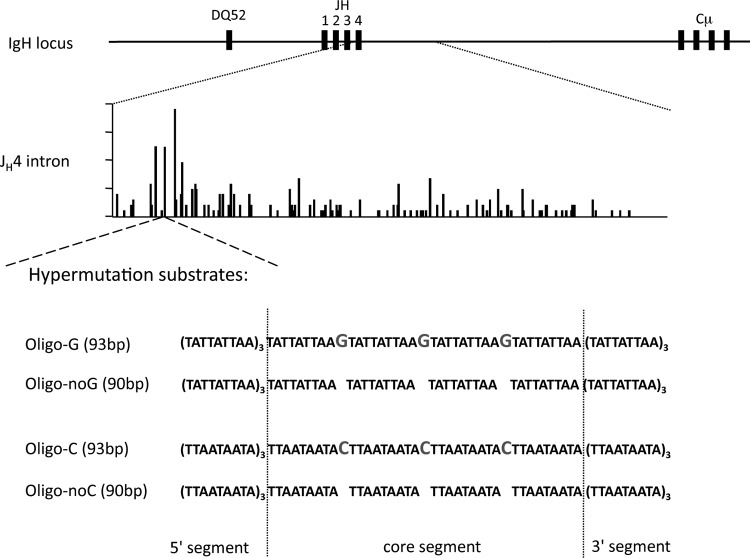
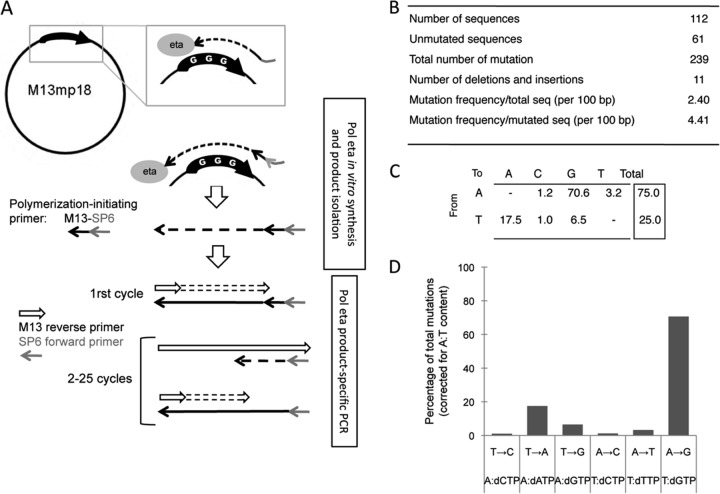
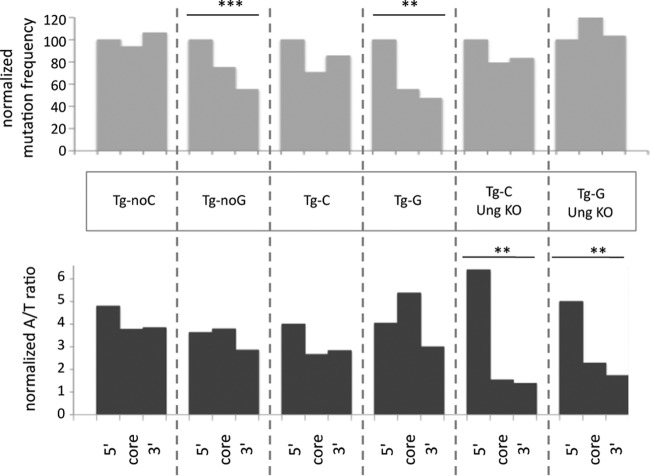
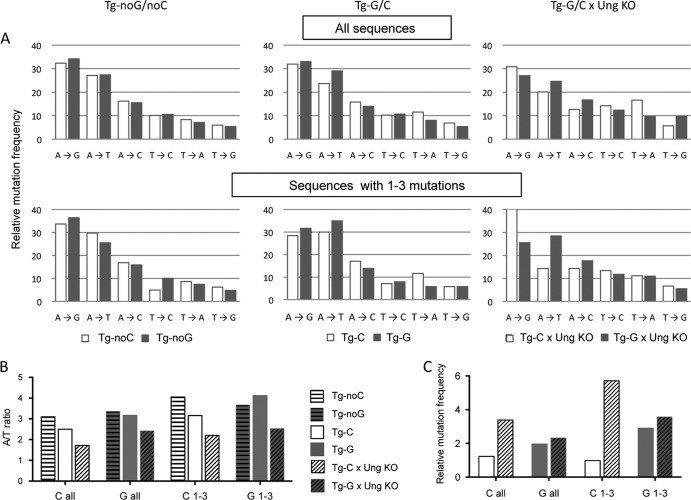

References
-
- Revy P, Muto T, Levy Y, Geissmann F, Plebani A, Sanal O, Catalan N, Forveille M, Dufourcq-Labelouse R, Gennery A, Tezcan I, Ersoy F, Kayserili H, Ugazio AG, Brousse N, Muramatsu M, Notarangelo LD, Kinoshita K, Honjo T, Fischer A, Durandy A. 2000. Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the hyper-IgM syndrome (HIGM2). Cell 102:565–575. 10.1016/S0092-8674(00)00079-9 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases