

Substrate-gated docking of pore subunit Tha4 in the TatC cavity initiates Tat translocase assembly
- PMID: 24711501
- PMCID: PMC3987133
- DOI: 10.1083/jcb.201311057
Substrate-gated docking of pore subunit Tha4 in the TatC cavity initiates Tat translocase assembly
Abstract
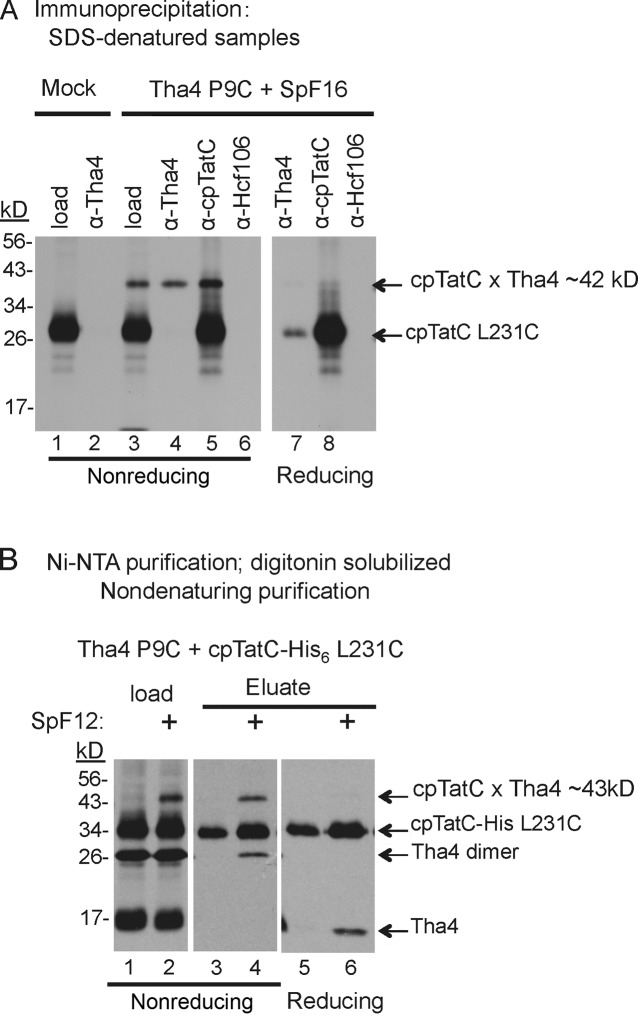
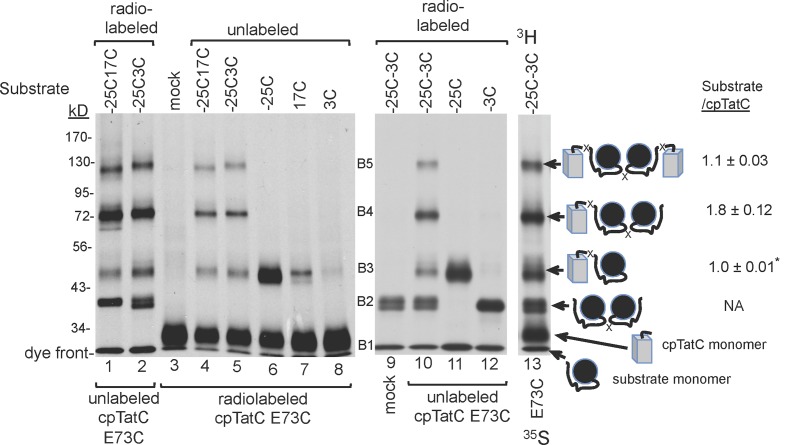
The twin-arginine translocase (Tat) transports folded proteins across tightly sealed membranes. cpTatC is the core component of the thylakoid translocase and coordinates transport through interactions with the substrate signal peptide and other Tat components, notably the Tha4 pore-forming component. Here, Cys-Cys matching mapped Tha4 contact sites on cpTatC and assessed the role of signal peptide binding on Tha4 assembly with the cpTatC-Hcf106 receptor complex. Tha4 made contact with a peripheral cpTatC site in nonstimulated membranes. In the translocase, Tha4 made an additional contact within the cup-shaped cavity of cpTatC that likely seeds Tha4 polymerization to form the pore. Substrate binding triggers assembly of Tha4 onto the interior site. We provide evidence that the substrate signal peptide inserts between cpTatC subunits arranged in a manner that conceivably forms an enclosed chamber. The location of the inserted signal peptide and the Tha4-cpTatC contact data suggest a model for signal peptide-gated Tha4 entry into the chamber to form the translocase.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
