

Casein kinase 2 phosphorylates GluA1 and regulates its surface expression
- PMID: 24712994
- PMCID: PMC4002167
- DOI: 10.1111/ejn.12494
Casein kinase 2 phosphorylates GluA1 and regulates its surface expression
Abstract
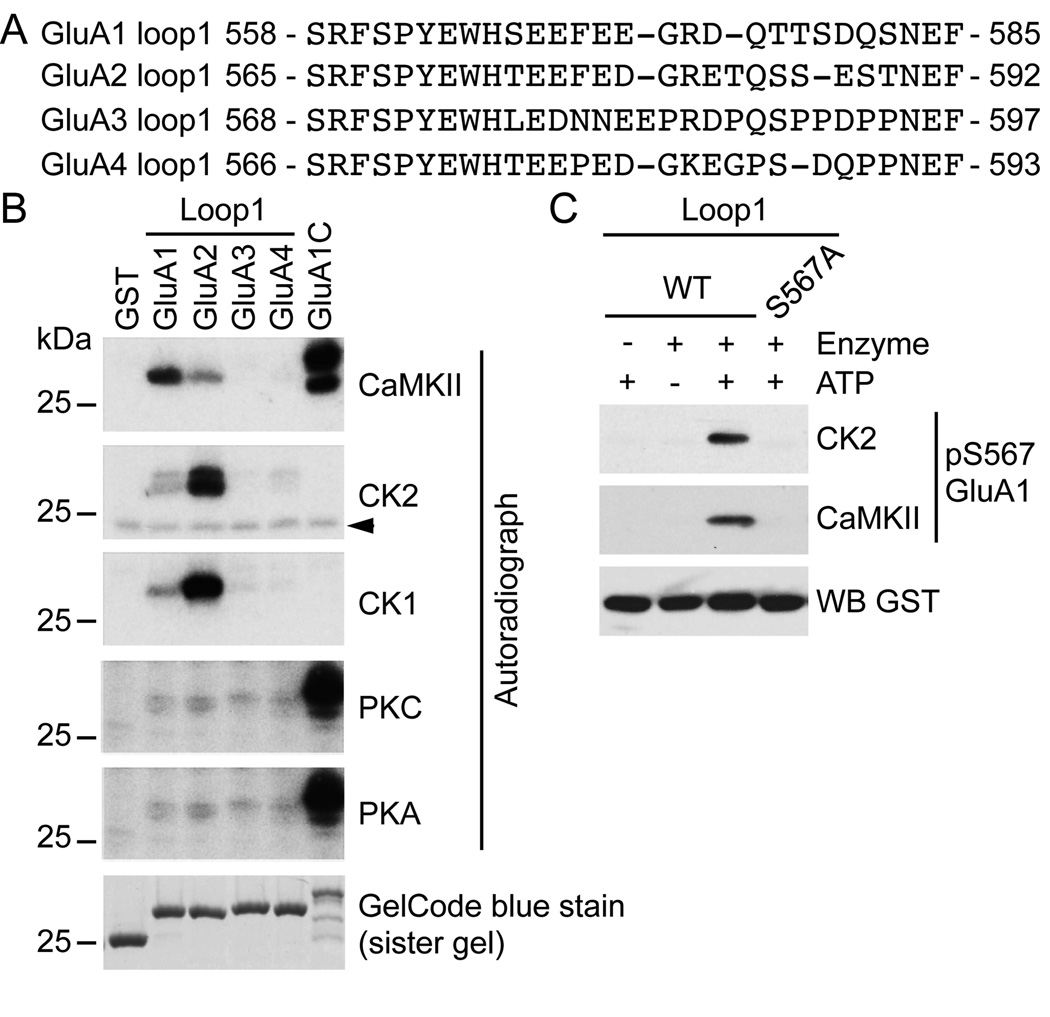
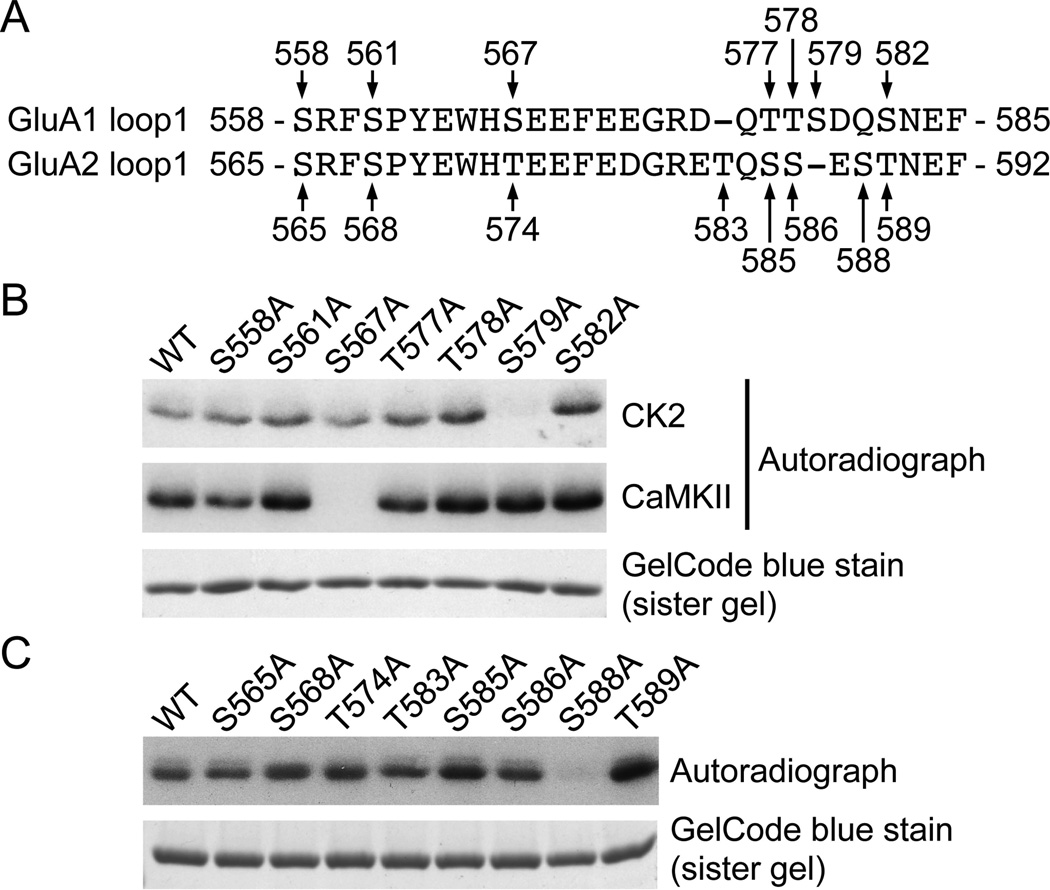
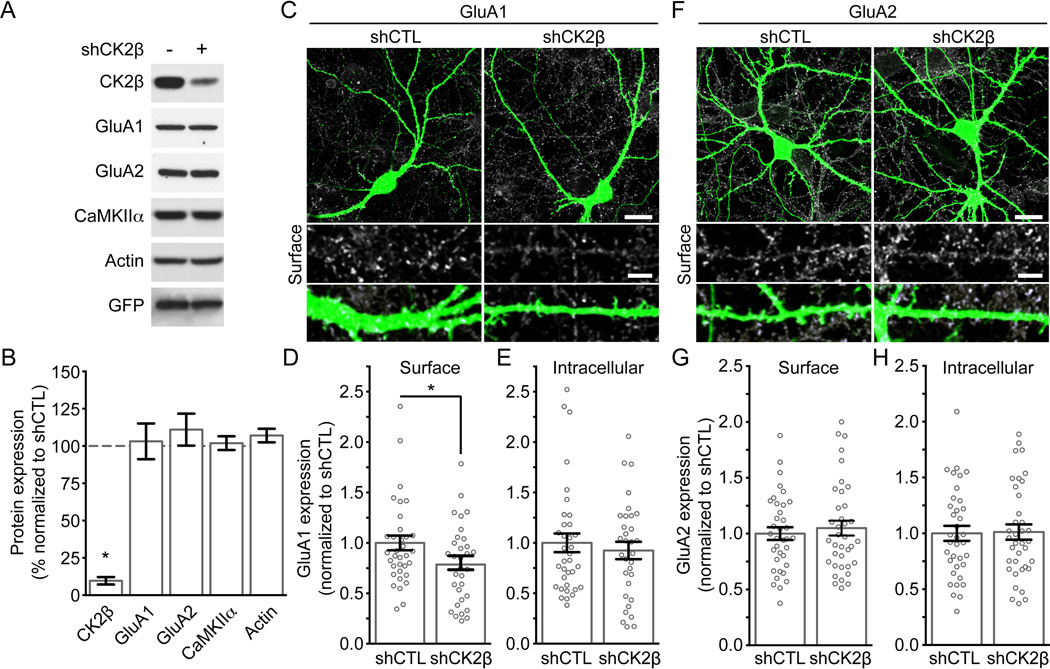
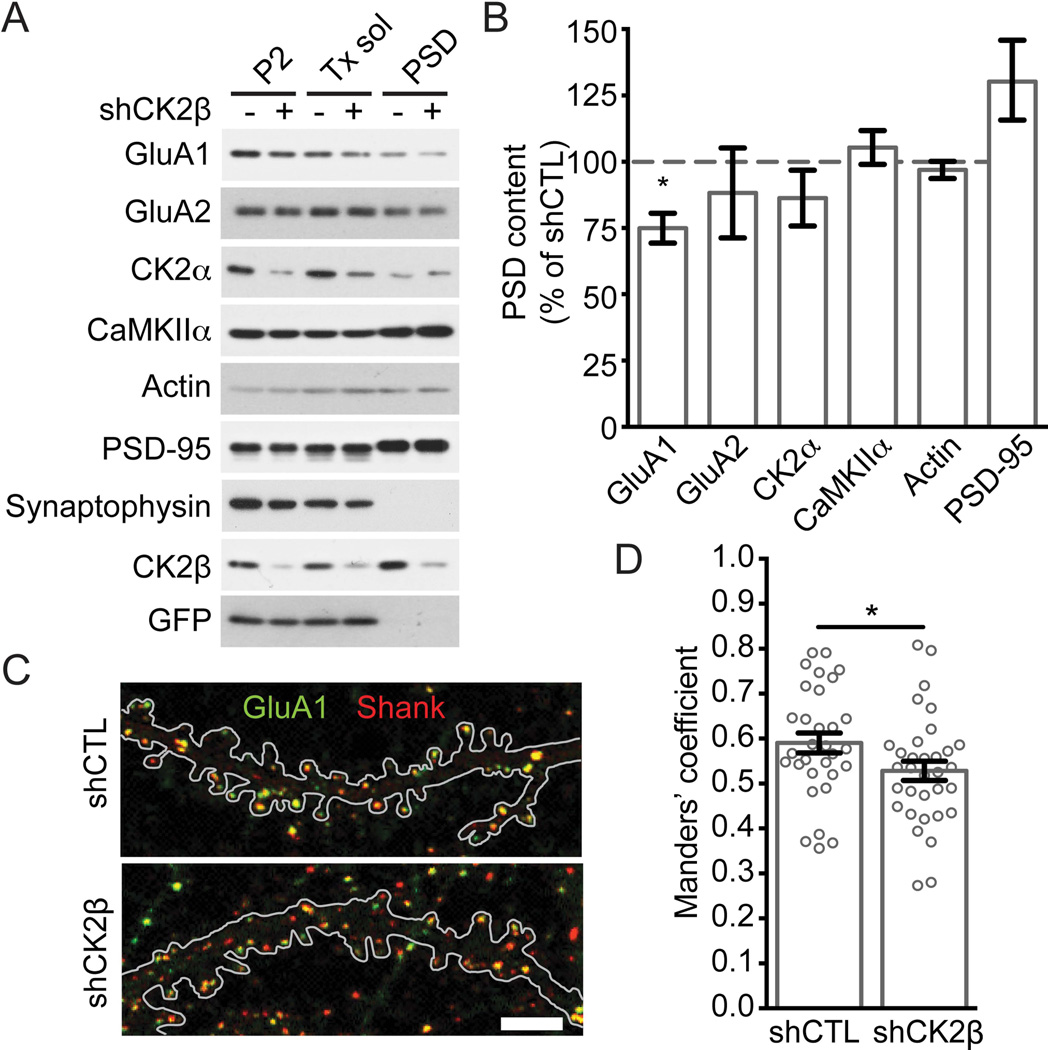
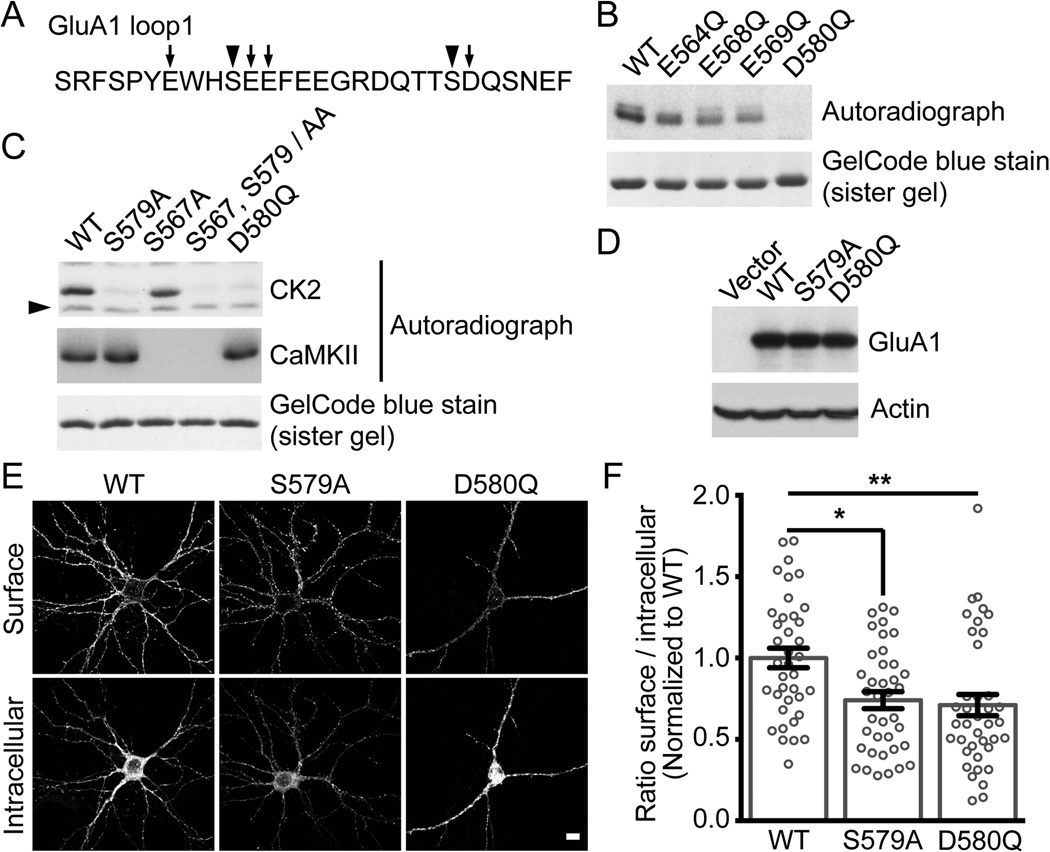
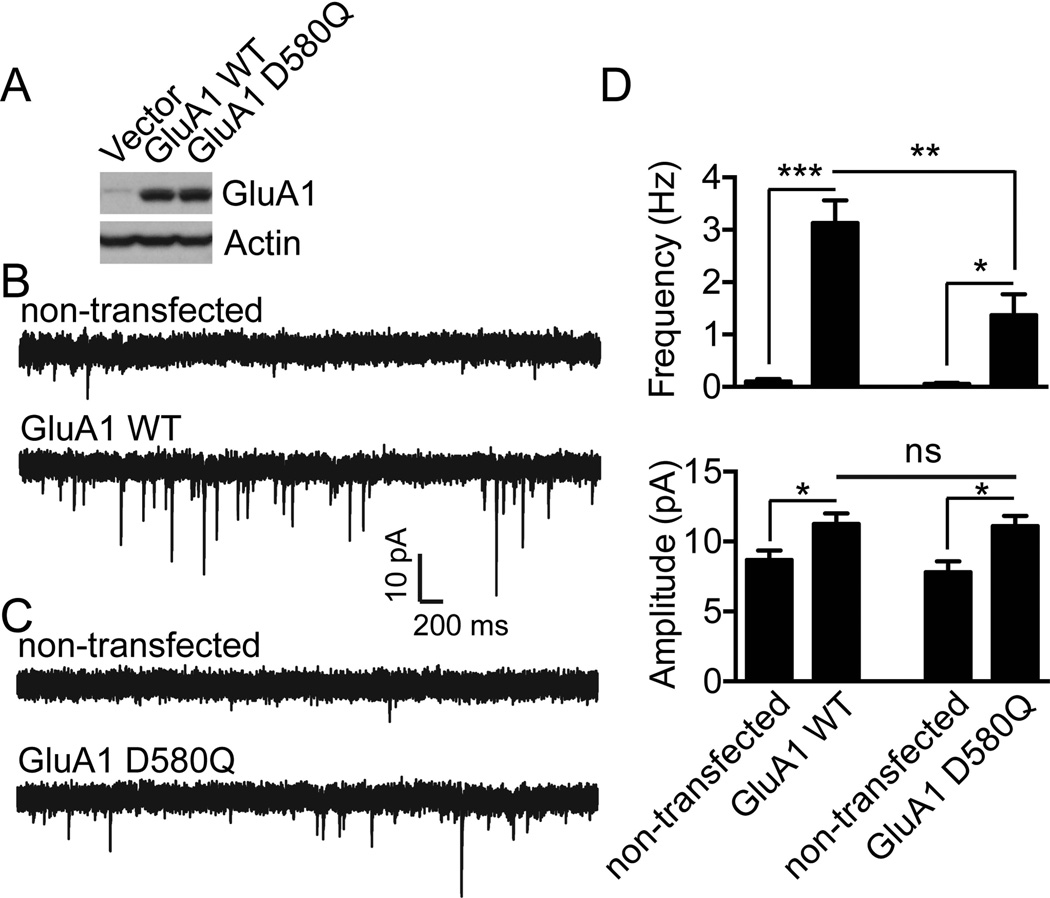
Controlling the density of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPARs) at synapses is essential for regulating the strength of excitatory neurotransmission. In particular, the phosphorylation of AMPARs is important for defining both synaptic expression and intracellular routing of receptors. Phosphorylation is a post-translational modification known to regulate many cellular events and the C-termini of glutamate receptors are important targets. Recently, the first intracellular loop1 region of the GluA1 subunit of AMPARs was reported to regulate synaptic targeting through phosphorylation of S567 by Ca2+ /calmodulin-dependent protein kinase II (CaMKII). Intriguingly, the loop1 region of all four AMPAR subunits contains many putative phosphorylation sites (S/T/Y), leaving the possibility that other kinases may regulate AMPAR surface expression via phosphorylation of the loop regions. To explore this hypothesis, we used in vitro phosphorylation assays with a small panel of purified kinases and found that casein kinase 2 (CK2) phosphorylates the GluA1 and GluA2 loop1 regions, but not GluA3 or GluA4. Interestingly, when we reduced the endogenous expression of CK2 using a specific short hairpin RNA against the regulatory subunit CK2β, we detected a reduction of GluA1 surface expression, whereas GluA2 was unchanged. Furthermore, we identified S579 of GluA1 as a substrate of CK2, and the expression of GluA1 phosphodeficient mutants in hippocampal neurons displayed reduced surface expression. Therefore, our study identifies CK2 as a regulator of GluA1 surface expression by phosphorylating the intracellular loop1 region.
Keywords: AMPAR; CK2; glutamate; synapse; trafficking.
Published 2014. This article is a U.S. Government work and is in the public domain in the USA.
Conflict of interest statement
All the authors declare no conflict of interest.
Figures
References
-
- Aksenova MV, Burbaeva GS, Kandror KV, Kapkov DV, Stepanov AS. The decreased level of casein kinase 2 in brain cortex of schizophrenic and Alzheimer’s disease patient. FEBS Lett. 1991;279:55–57. - PubMed
-
- Barria A, Derkach V, Soderling T. Identification of the Ca2+/calmodulin-dependent protein kinase II regulatory phosphorylation site in the alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate-type glutamate receptor. J. Biol. Chem. 1997;272:32727–32730. - PubMed
-
- Bellone C, Nicoll RA. Rapid bidirectional switching of synaptic NMDA receptors. Neuron. 2007;55:779–785. - PubMed
-
- Belmeguenai A, Hosy E, Bengtsson F, Pedroarena CM, Piochon C, Teuling E, He Q, Ohtsuki G, De Jeu MT, Elgersma Y, De Zeeuw CI, Jörntell H, Hansel C. Intrinsic plasticity complements long-term potentiation in parallel fiber input gain control in cerebellar Purkinje cells. J. Neurosci. 2010;30:13630–13643. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
