

Structure and mechanism of DNA polymerase β
- PMID: 24717170
- PMCID: PMC4018062
- DOI: 10.1021/bi500139h
Structure and mechanism of DNA polymerase β
Abstract
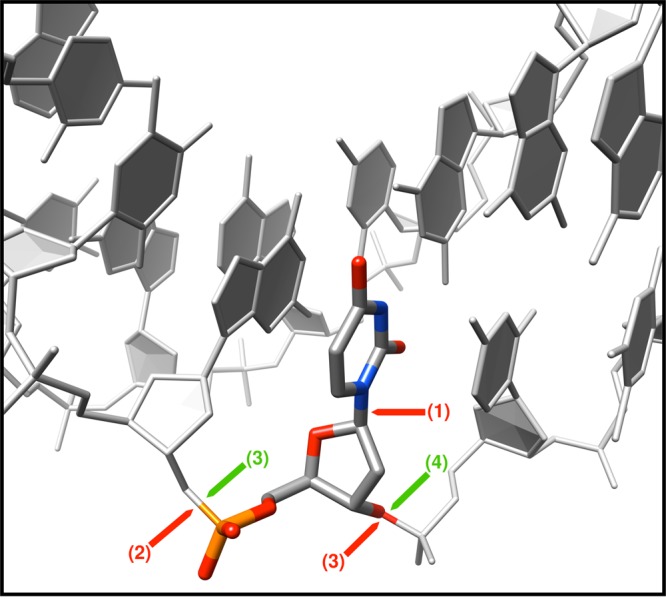
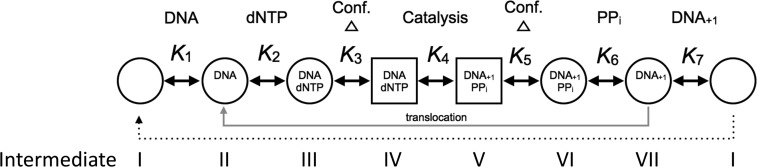
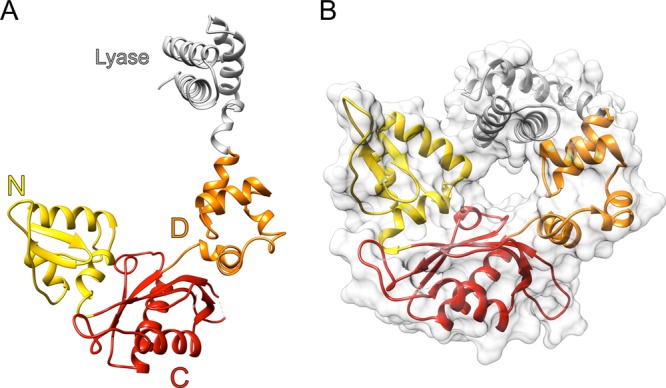
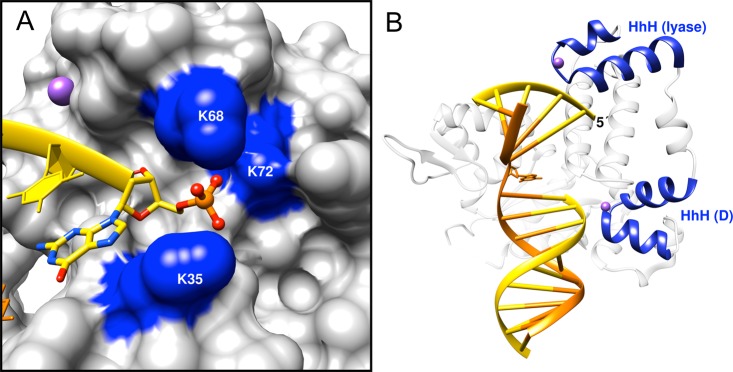
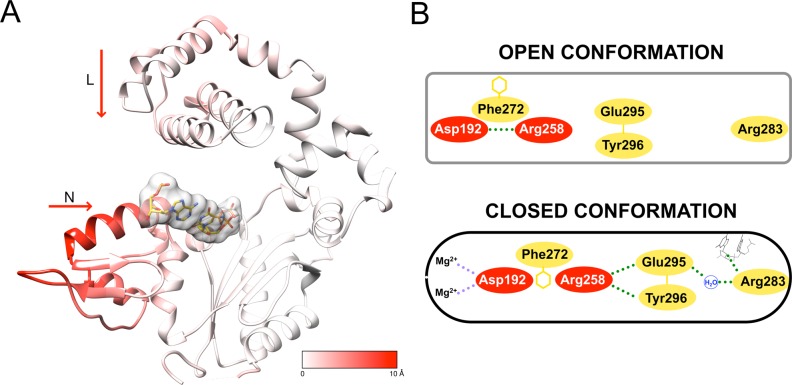
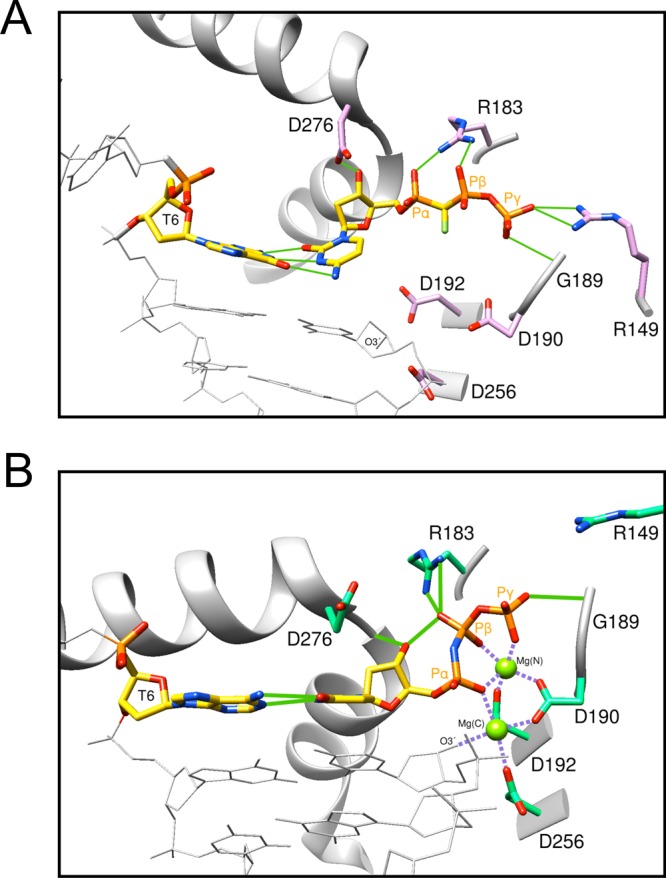
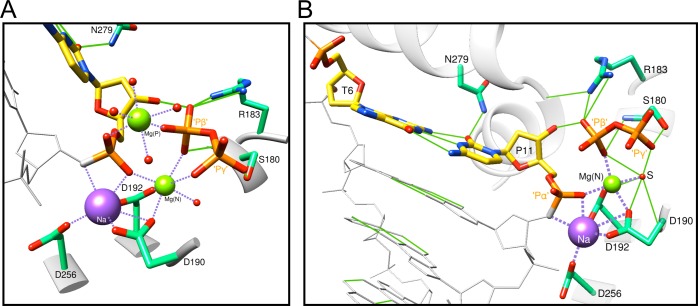
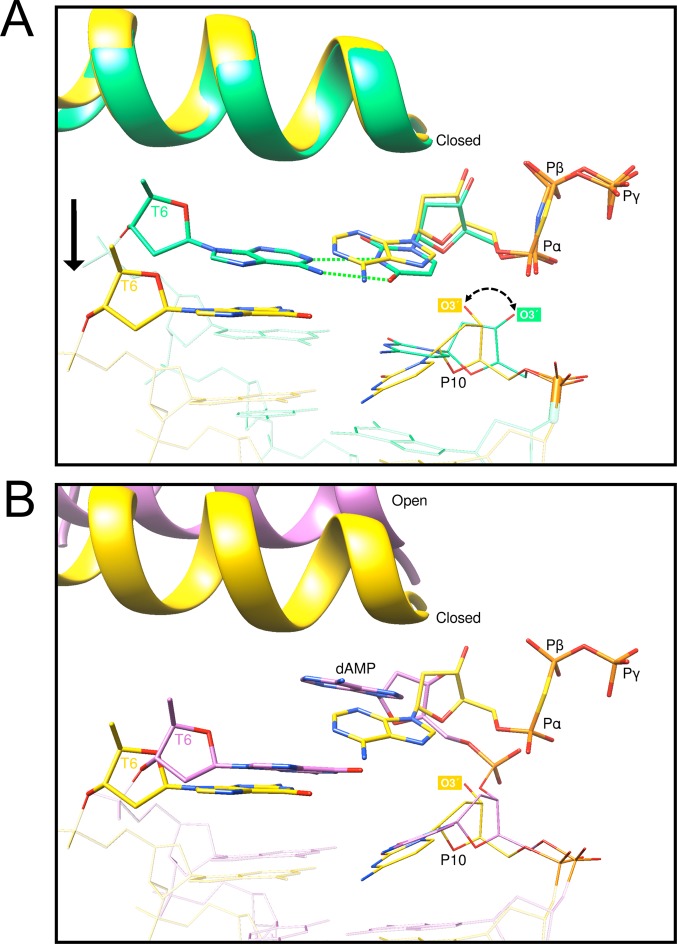
DNA polymerase (pol) β is a small eukaryotic DNA polymerase composed of two domains. Each domain contributes an enzymatic activity (DNA synthesis and deoxyribose phosphate lyase) during the repair of simple base lesions. These domains are termed the polymerase and lyase domains, respectively. Pol β has been an excellent model enzyme for studying the nucleotidyl transferase reaction and substrate discrimination at a molecular level. In this review, recent crystallographic studies of pol β in various liganded and conformational states during the insertion of right and wrong nucleotides as well as during the bypass of damaged DNA (apurinic sites and 8-oxoguanine) are described. Structures of these catalytic intermediates provide unexpected insights into mechanisms by which DNA polymerases enhance genome stability. These structures also provide an improved framework that permits computational studies to facilitate the interpretation of detailed kinetic analyses of this model enzyme.
Figures
References
-
- Beard W. A.; Wilson S. H. (2000) Structural design of a eukaryotic DNA repair polymerase: DNA polymerase β. Mutat. Res. 460, 231–244. - PubMed
-
- Beard W. A.; Wilson S. H. (2006) Structure and mechanism of DNA polymerase β. Chem. Rev. 106, 361–382. - PubMed
-
- Nakamura J.; Swenberg J. A. (1999) Endogenous apurinic/apyrimidinic sites in genomic DNA of mammalian tissues. Cancer Res. 59, 2522–2526. - PubMed
-
- Lindahl T.; Nyberg B. (1972) Rate of depurination of native deoxyribonucleic acid. Biochemistry 11, 3610–3618. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
