

Epidermal Merkel cells are mechanosensory cells that tune mammalian touch receptors
- PMID: 24717432
- PMCID: PMC4097312
- DOI: 10.1038/nature13250
Epidermal Merkel cells are mechanosensory cells that tune mammalian touch receptors
Abstract
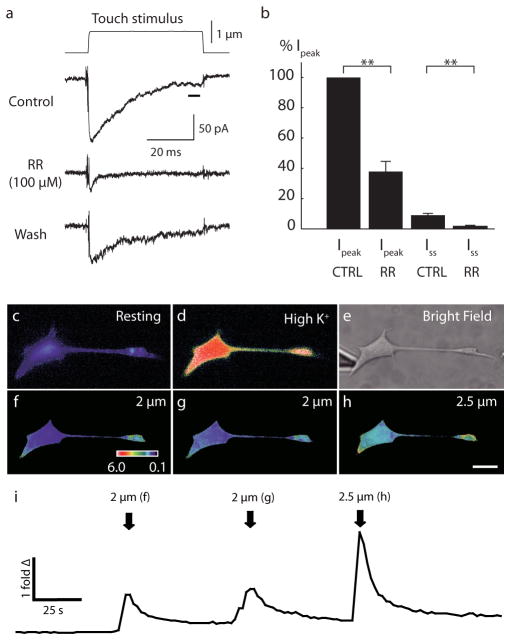
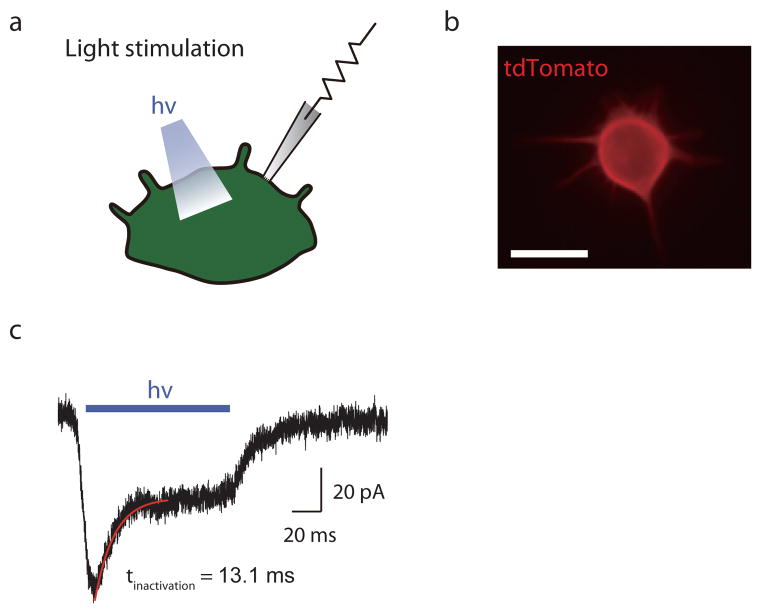
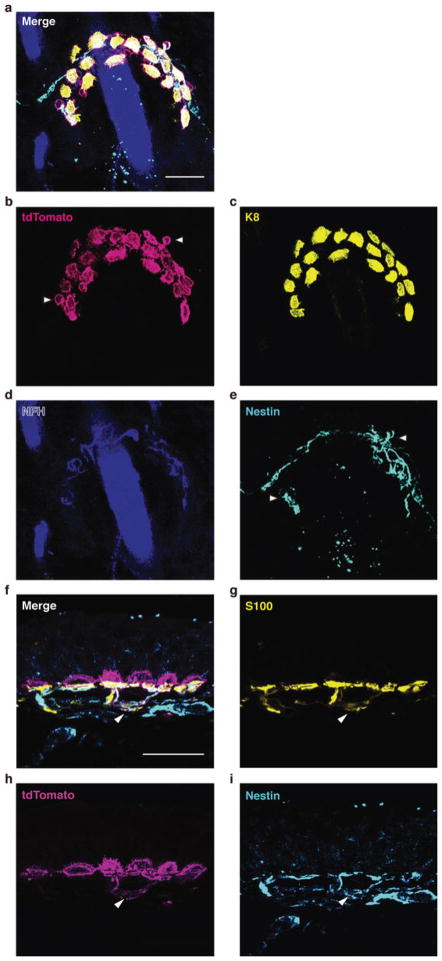
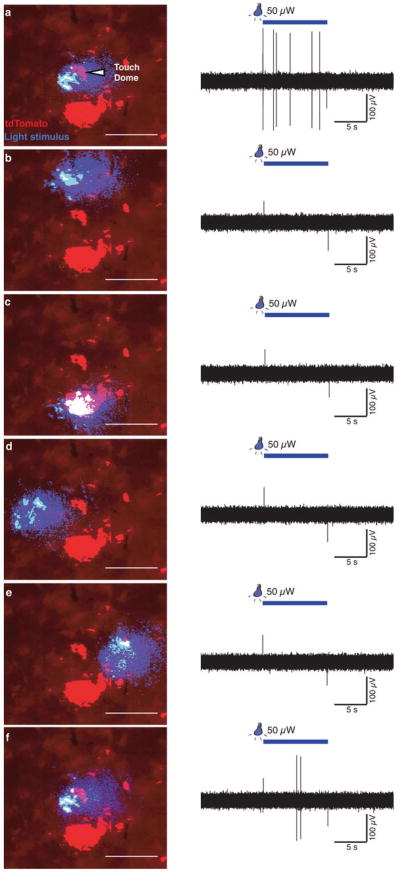
Touch submodalities, such as flutter and pressure, are mediated by somatosensory afferents whose terminal specializations extract tactile features and encode them as action potential trains with unique activity patterns. Whether non-neuronal cells tune touch receptors through active or passive mechanisms is debated. Terminal specializations are thought to function as passive mechanical filters analogous to the cochlea's basilar membrane, which deconstructs complex sounds into tones that are transduced by mechanosensory hair cells. The model that cutaneous specializations are merely passive has been recently challenged because epidermal cells express sensory ion channels and neurotransmitters; however, direct evidence that epidermal cells excite tactile afferents is lacking. Epidermal Merkel cells display features of sensory receptor cells and make 'synapse-like' contacts with slowly adapting type I (SAI) afferents. These complexes, which encode spatial features such as edges and texture, localize to skin regions with high tactile acuity, including whisker follicles, fingertips and touch domes. Here we show that Merkel cells actively participate in touch reception in mice. Merkel cells display fast, touch-evoked mechanotransduction currents. Optogenetic approaches in intact skin show that Merkel cells are both necessary and sufficient for sustained action-potential firing in tactile afferents. Recordings from touch-dome afferents lacking Merkel cells demonstrate that Merkel cells confer high-frequency responses to dynamic stimuli and enable sustained firing. These data are the first, to our knowledge, to directly demonstrate a functional, excitatory connection between epidermal cells and sensory neurons. Together, these findings indicate that Merkel cells actively tune mechanosensory responses to facilitate high spatio-temporal acuity. Moreover, our results indicate a division of labour in the Merkel cell-neurite complex: Merkel cells signal static stimuli, such as pressure, whereas sensory afferents transduce dynamic stimuli, such as moving gratings. Thus, the Merkel cell-neurite complex is an unique sensory structure composed of two different receptor cell types specialized for distinct elements of discriminative touch.
Figures
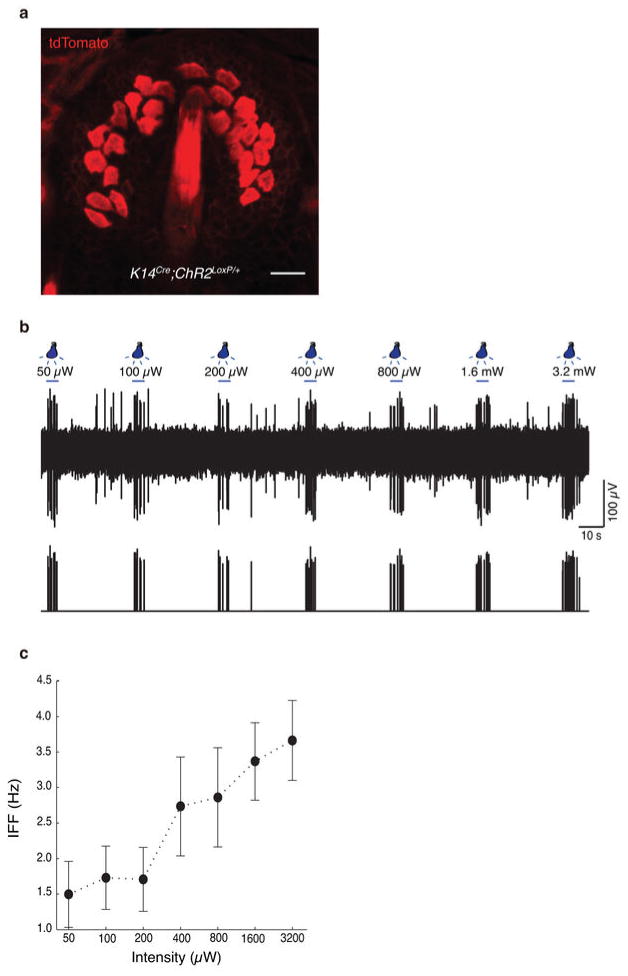
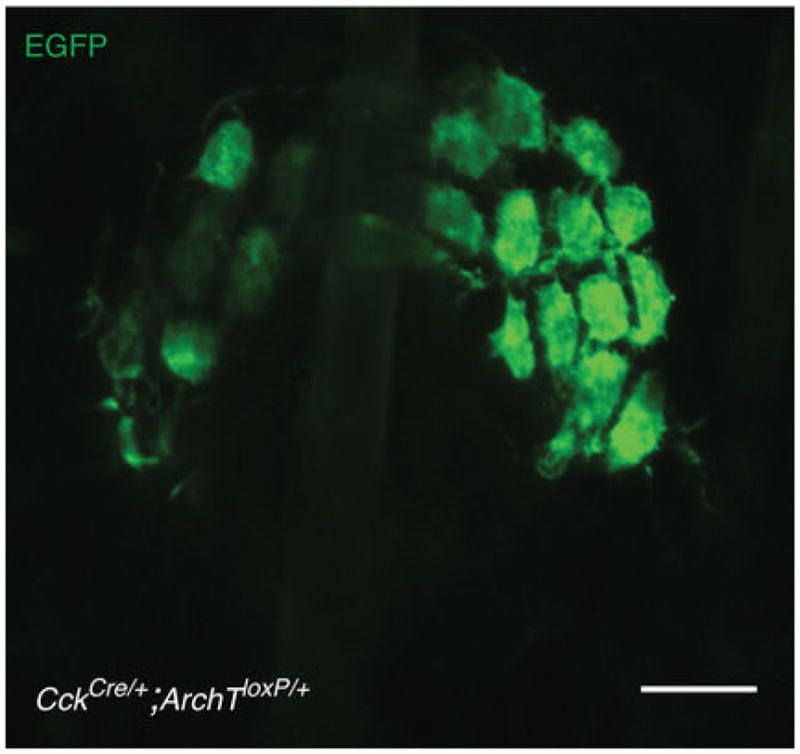
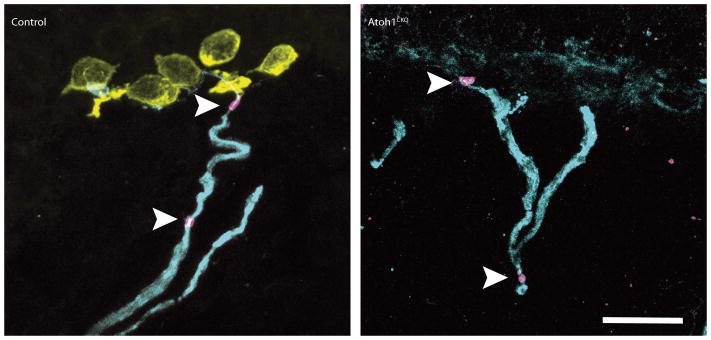
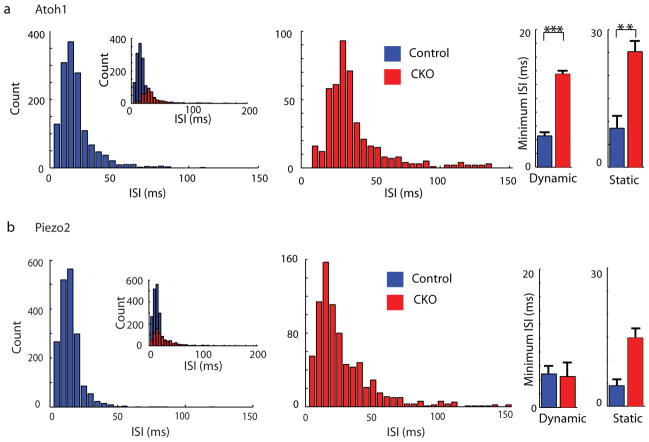
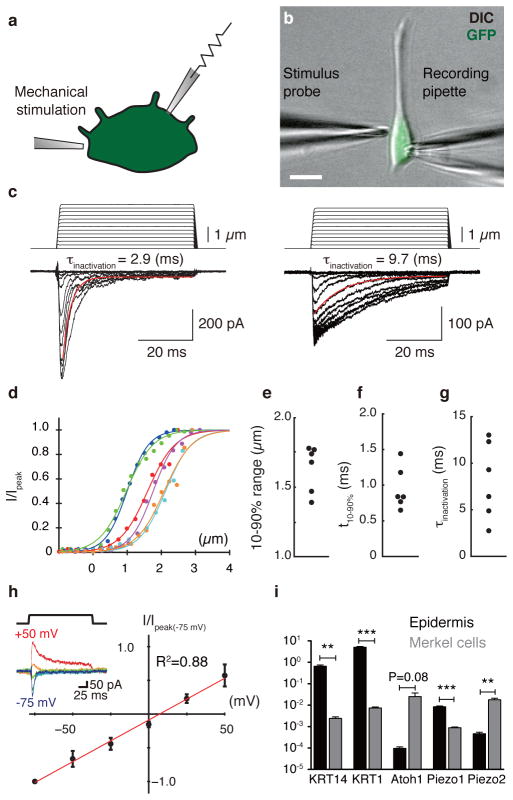
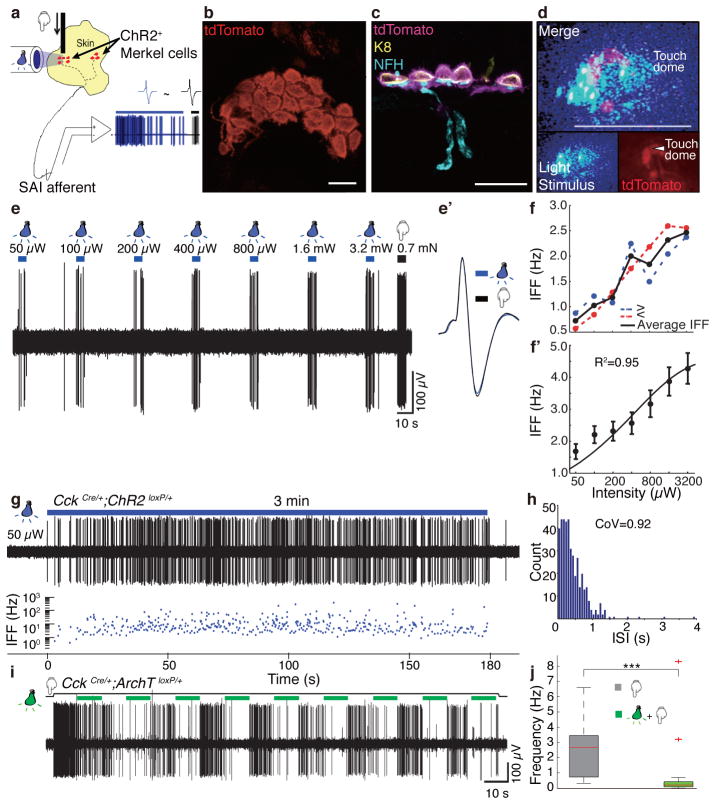
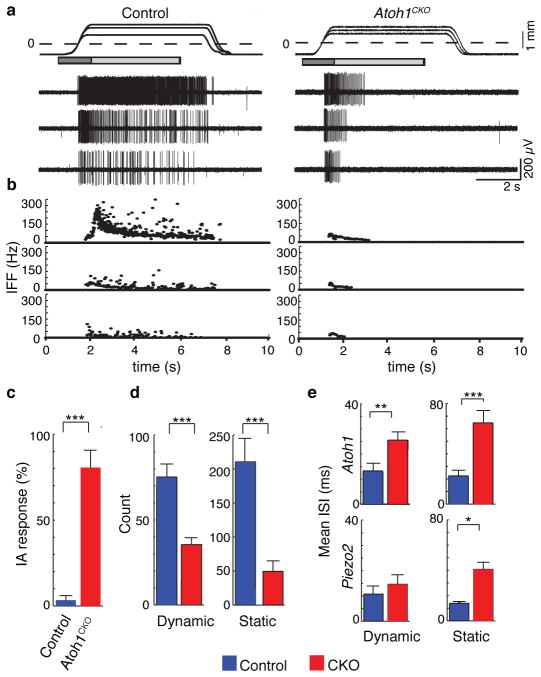
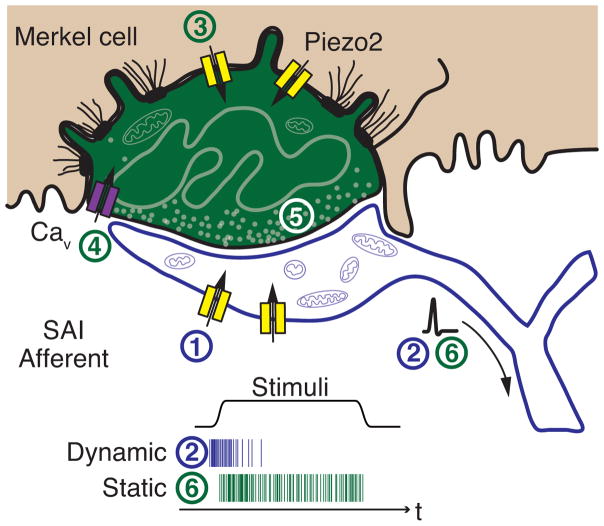
Comment in
-
Sensory transduction: Merkel cells bring a delicate touch.Nat Rev Neurosci. 2014 Jun;15(6):348-9. doi: 10.1038/nrn3748. Epub 2014 Apr 25. Nat Rev Neurosci. 2014. PMID: 24762930 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
- 5T32HL087745-05/HL/NHLBI NIH HHS/United States
- R01 DE022358/DE/NIDCR NIH HHS/United States
- P30 CA125123/CA/NCI NIH HHS/United States
- P30 CA013696/CA/NCI NIH HHS/United States
- R21 AR062307/AR/NIAMS NIH HHS/United States
- R01DE022358/DE/NIDCR NIH HHS/United States
- P30 AR044535/AR/NIAMS NIH HHS/United States
- F32 NS080544/NS/NINDS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01AR051219/AR/NIAMS NIH HHS/United States
- R21AR062307/AR/NIAMS NIH HHS/United States
- P30CA013696/CA/NCI NIH HHS/United States
- P30CA125123/CA/NCI NIH HHS/United States
- P30AR044535/AR/NIAMS NIH HHS/United States
- R01 AR051219/AR/NIAMS NIH HHS/United States
- F32NS080544/NS/NINDS NIH HHS/United States
- T32 HL087745/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
