

Recapitulation of morphogenetic cell shape changes enables wound re-epithelialisation
- PMID: 24718989
- PMCID: PMC3994776
- DOI: 10.1242/dev.107045
Recapitulation of morphogenetic cell shape changes enables wound re-epithelialisation
Abstract
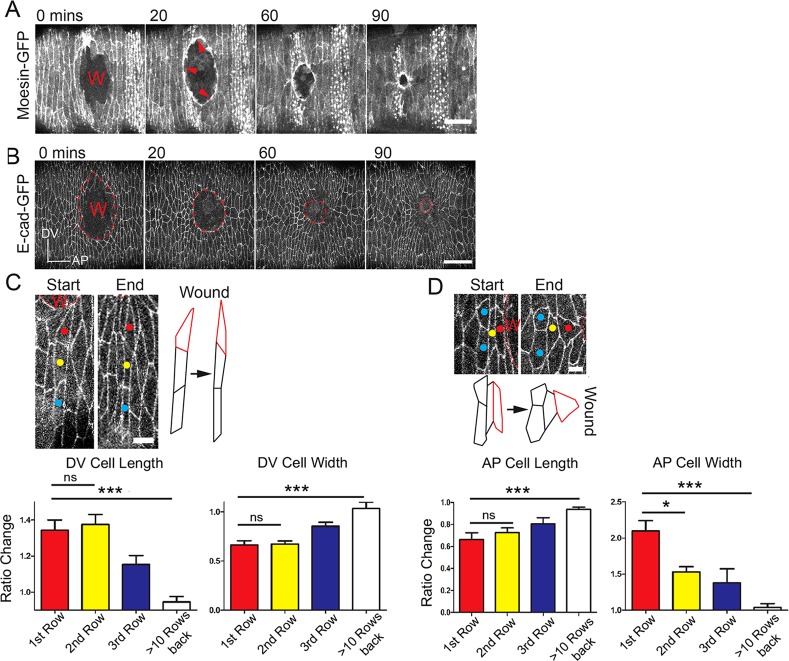
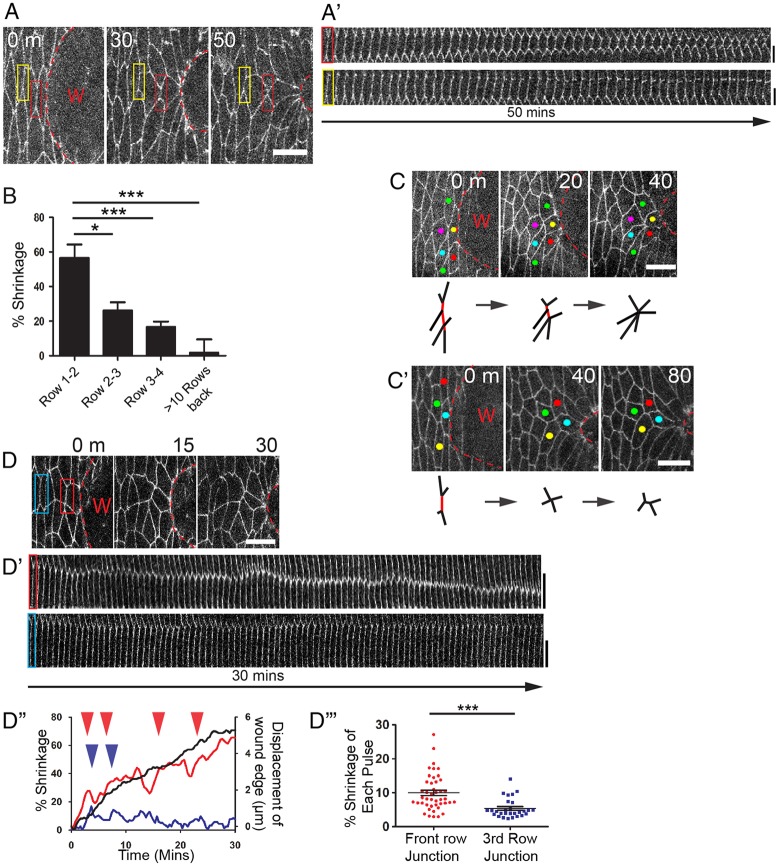
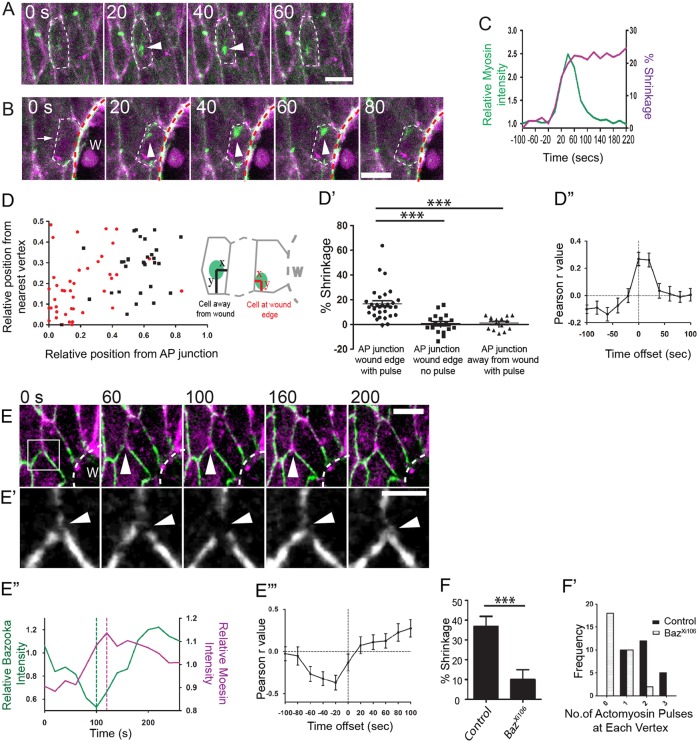
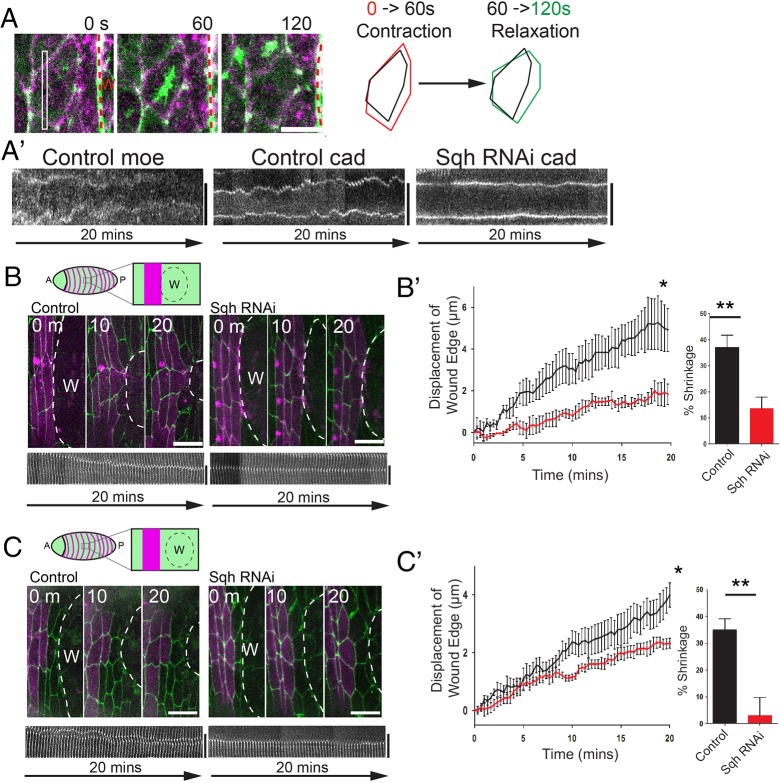
Wound repair is a fundamental, conserved mechanism for maintaining tissue homeostasis and shares many parallels with embryonic morphogenesis. Small wounds in simple epithelia rapidly assemble a contractile actomyosin cable at their leading edge, as well as dynamic filopodia that finally knit the wound edges together. Most studies of wound re-epithelialisation have focused on the actin machineries that assemble in the leading edge of front row cells and that resemble the contractile mechanisms that drive morphogenetic episodes, including Drosophila dorsal closure, but, clearly, multiple cell rows back must also contribute for efficient repair of the wound. Here, we examine the role of cells back from the wound edge and show that they also stretch towards the wound and cells anterior-posterior to the wound edge rearrange their junctions with neighbours to drive cell intercalation events. This process in anterior-posterior cells is active and dependent on pulses of actomyosin that lead to ratcheted shrinkage of junctions; the actomyosin pulses are targeted to breaks in the cell polarity protein Par3 at cell vertices. Inhibiting actomyosin dynamics back from the leading edge prevents junction shrinkage and inhibits the wound edge from advancing. These events recapitulate cell rearrangements that occur during germband extension, in which intercalation events drive the elongation of tissues.
Keywords: Actomyosin behaviour; Drosophila embryo; Wound healing.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
