

Neutrophils recruited by IL-22 in peripheral tissues function as TRAIL-dependent antiviral effectors against MCMV
- PMID: 24721575
- PMCID: PMC3989063
- DOI: 10.1016/j.chom.2014.03.003
Neutrophils recruited by IL-22 in peripheral tissues function as TRAIL-dependent antiviral effectors against MCMV
Abstract
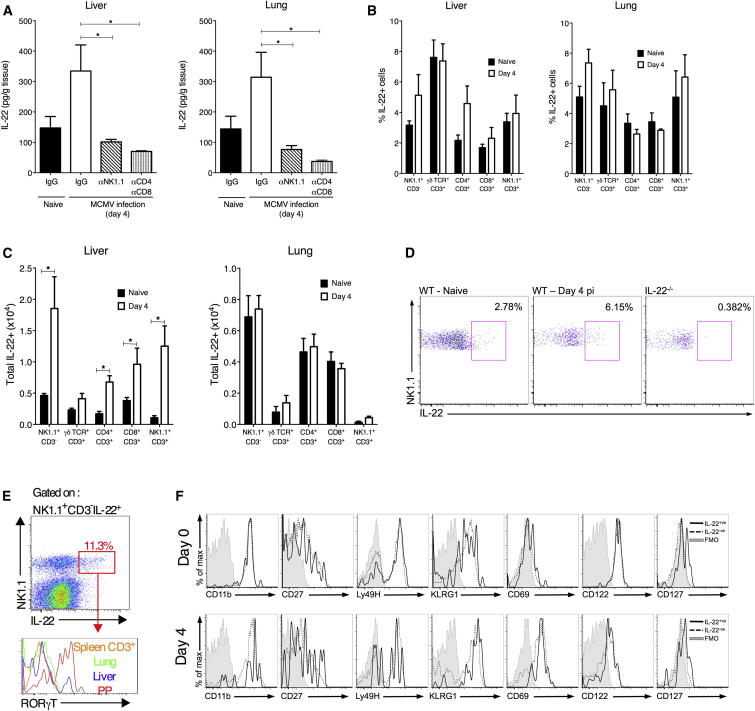
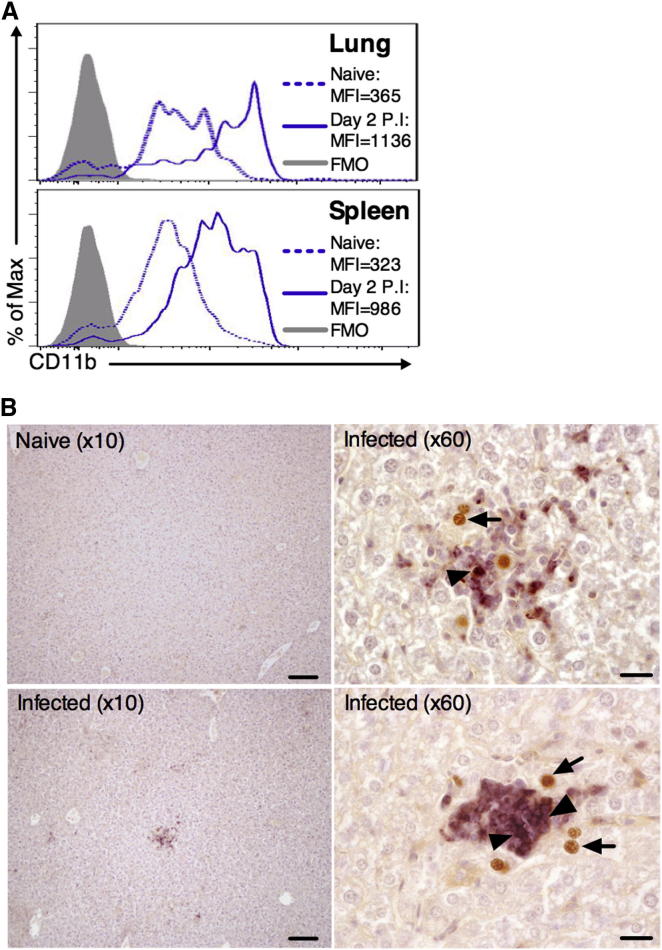
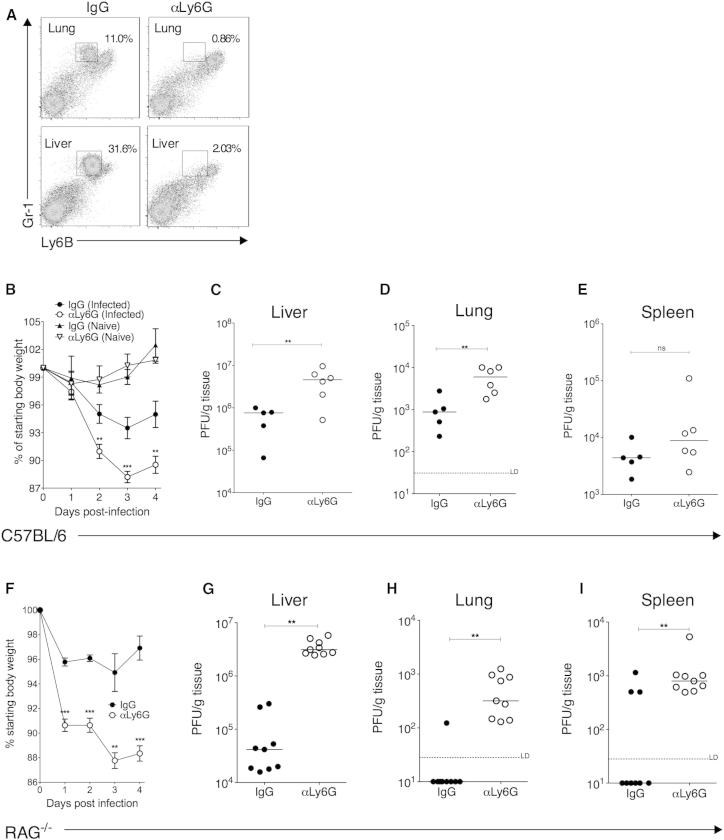
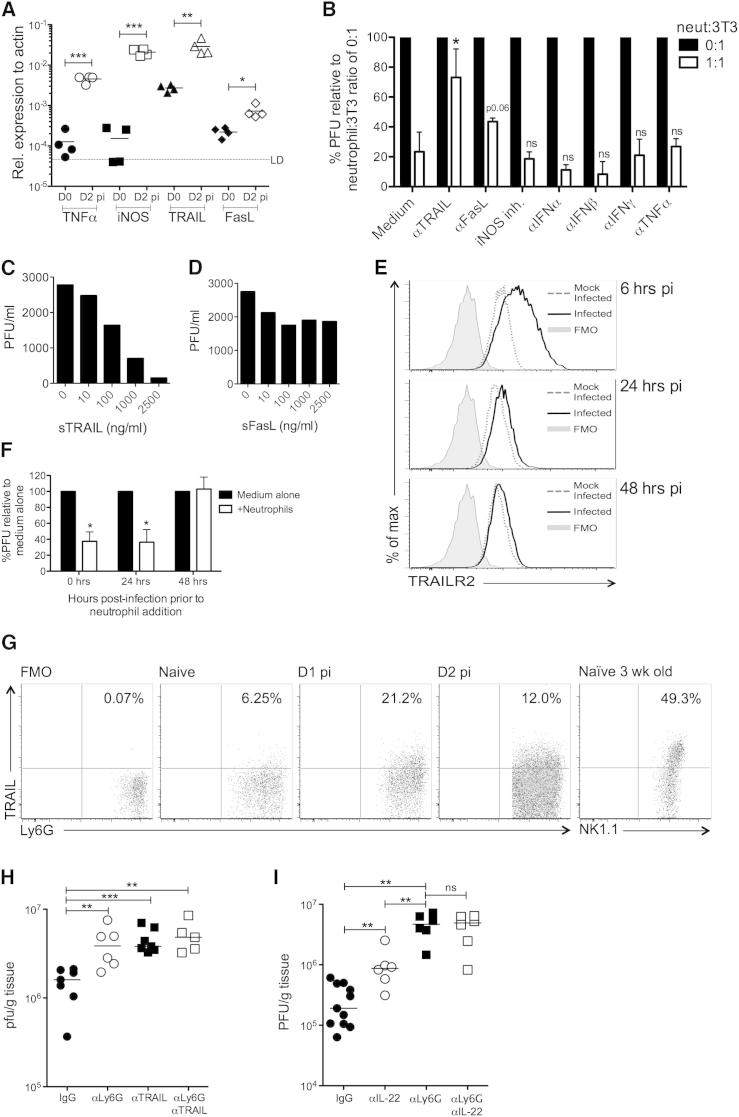
During primary infection, murine cytomegalovirus (MCMV) spreads systemically, resulting in virus replication and pathology in multiple organs. This disseminated infection is ultimately controlled, but the underlying immune defense mechanisms are unclear. Investigating the role of the cytokine IL-22 in MCMV infection, we discovered an unanticipated function for neutrophils as potent antiviral effector cells that restrict viral replication and associated pathogenesis in peripheral organs. NK-, NKT-, and T cell-secreted IL-22 orchestrated antiviral neutrophil-mediated responses via induction in stromal nonhematopoietic tissue of the neutrophil-recruiting chemokine CXCL1. The antiviral effector properties of infiltrating neutrophils were directly linked to the expression of TNF-related apoptosis-inducing ligand (TRAIL). Our data identify a role for neutrophils in antiviral defense, and establish a functional link between IL-22 and the control of antiviral neutrophil responses that prevents pathogenic herpesvirus infection in peripheral organs.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Andoniou C.E., van Dommelen S.L., Voigt V., Andrews D.M., Brizard G., Asselin-Paturel C., Delale T., Stacey K.J., Trinchieri G., Degli-Esposti M.A. Interaction between conventional dendritic cells and natural killer cells is integral to the activation of effective antiviral immunity. Nat. Immunol. 2005;6:1011–1019. - PubMed
-
- Andrews D.M., Scalzo A.A., Yokoyama W.M., Smyth M.J., Degli-Esposti M.A. Functional interactions between dendritic cells and NK cells during viral infection. Nat. Immunol. 2003;4:175–181. - PubMed
-
- Banks T.A., Rickert S., Benedict C.A., Ma L., Ko M., Meier J., Ha W., Schneider K., Granger S.W., Turovskaya O. A lymphotoxin-IFN-beta axis essential for lymphocyte survival revealed during cytomegalovirus infection. J. Immunol. 2005;174:7217–7225. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI101423/AI/NIAID NIH HHS/United States
- G0601617/MRC_/Medical Research Council/United Kingdom
- G1000236/MRC_/Medical Research Council/United Kingdom
- WT090323MA/WT_/Wellcome Trust/United Kingdom
- R56 AI101423/AI/NIAID NIH HHS/United States
- G0700142/MRC_/Medical Research Council/United Kingdom
- BB/F009836/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/L018373/1/MRC_/Medical Research Council/United Kingdom
- 091663/WT_/Wellcome Trust/United Kingdom
- AI101423/AI/NIAID NIH HHS/United States
- MR/J002151/1/MRC_/Medical Research Council/United Kingdom
- WT098026MA/WT_/Wellcome Trust/United Kingdom
- BBF0098361/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
