

Renal-retinal ciliopathy gene Sdccag8 regulates DNA damage response signaling
- PMID: 24722439
- PMCID: PMC4214515
- DOI: 10.1681/ASN.2013050565
Renal-retinal ciliopathy gene Sdccag8 regulates DNA damage response signaling
Abstract
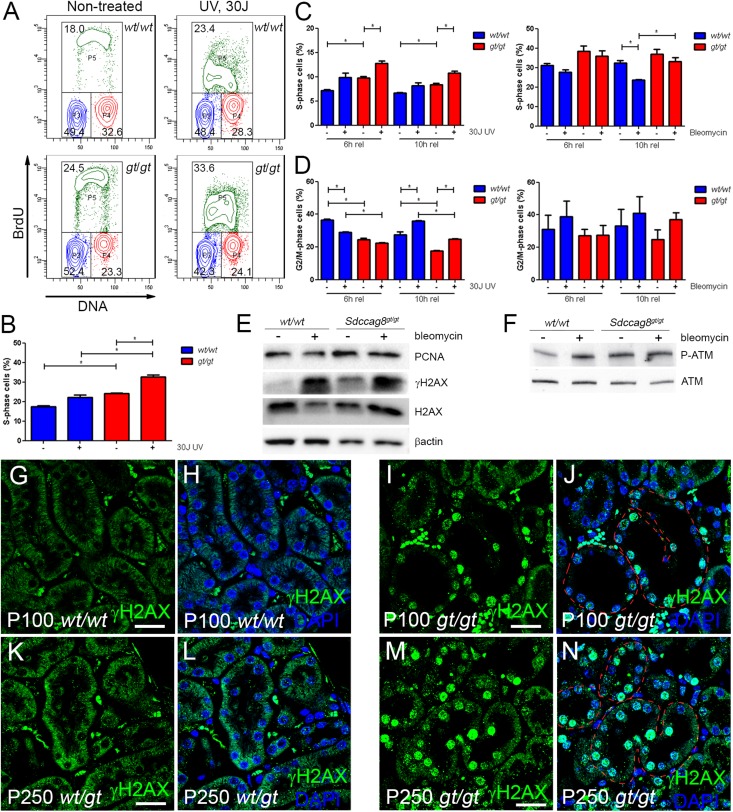
Nephronophthisis-related ciliopathies (NPHP-RCs) are developmental and degenerative kidney diseases that are frequently associated with extrarenal pathologies such as retinal degeneration, obesity, and intellectual disability. We recently identified mutations in a gene encoding the centrosomal protein SDCCAG8 as causing NPHP type 10 in humans. To study the role of Sdccag8 in disease pathogenesis, we generated a Sdccag8 gene-trap mouse line. Homozygous Sdccag8(gt/gt) mice lacked the wild-type Sdccag8 transcript and protein, and recapitulated the human phenotypes of NPHP and retinal degeneration. These mice exhibited early onset retinal degeneration that was associated with rhodopsin mislocalization in the photoreceptors and reduced cone cell numbers, and led to progressive loss of vision. By contrast, renal histologic changes occurred later, and no global ciliary defects were observed in the kidneys. Instead, renal pathology was associated with elevated levels of DNA damage response signaling activity. Cell culture studies confirmed the aberrant activation of DNA damage response in Sdccag8(gt/gt)-derived cells, characterized by elevated levels of γH2AX and phosphorylated ATM and cell cycle profile abnormalities. Our analysis of Sdccag8(gt/gt) mice indicates that the pleiotropic phenotypes in these mice may arise through multiple tissue-specific disease mechanisms.
Copyright © 2014 by the American Society of Nephrology.
Figures
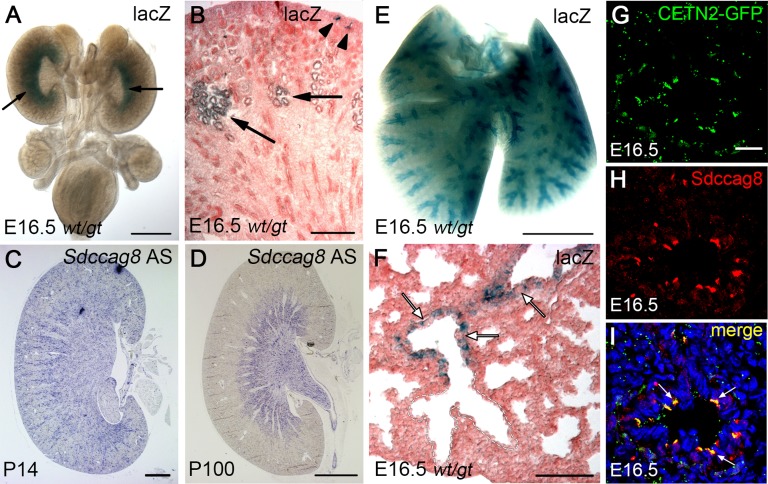
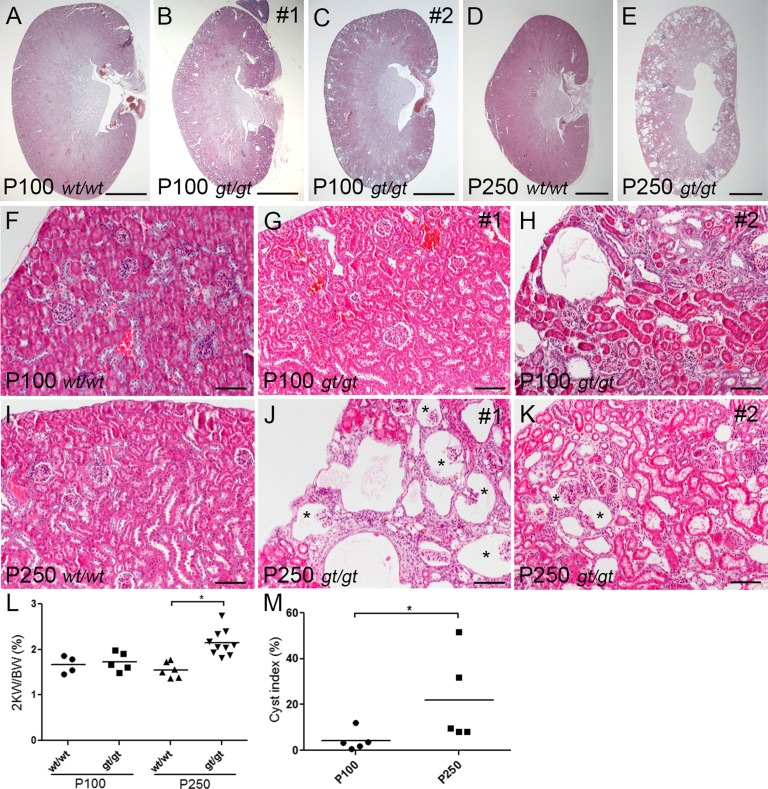
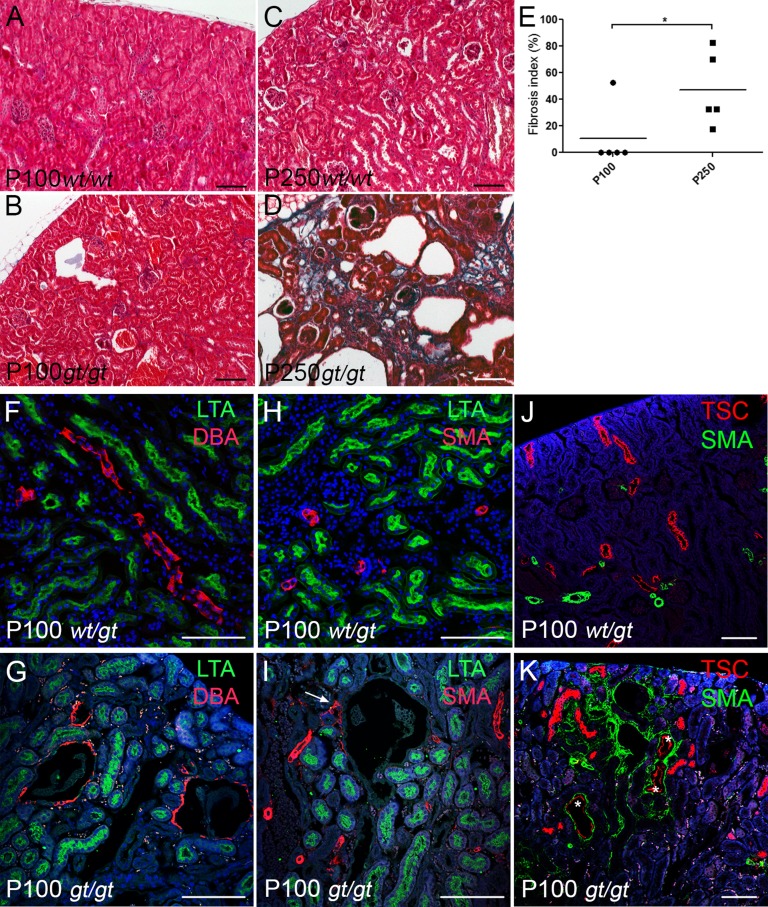
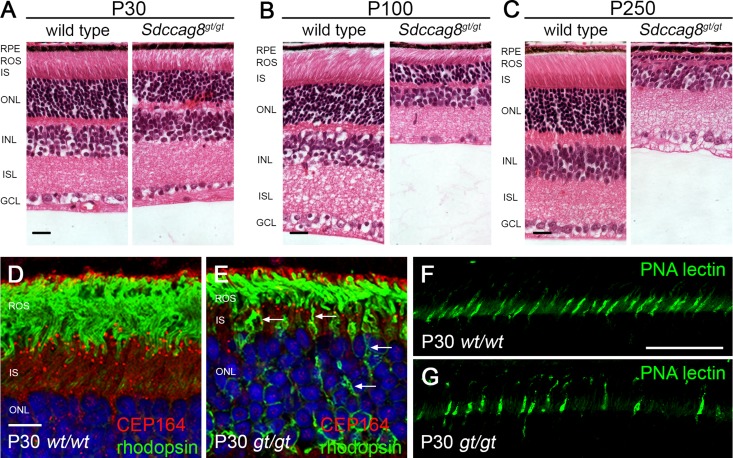
References
-
- Otto EA, Hurd TW, Airik R, Chaki M, Zhou W, Stoetzel C, Patil SB, Levy S, Ghosh AK, Murga-Zamalloa CA, van Reeuwijk J, Letteboer SJ, Sang L, Giles RH, Liu Q, Coene KL, Estrada-Cuzcano A, Collin RW, McLaughlin HM, Held S, Kasanuki JM, Ramaswami G, Conte J, Lopez I, Washburn J, Macdonald J, Hu J, Yamashita Y, Maher ER, Guay-Woodford LM, Neumann HP, Obermüller N, Koenekoop RK, Bergmann C, Bei X, Lewis RA, Katsanis N, Lopes V, Williams DS, Lyons RH, Dang CV, Brito DA, Dias MB, Zhang X, Cavalcoli JD, Nürnberg G, Nürnberg P, Pierce EA, Jackson PK, Antignac C, Saunier S, Roepman R, Dollfus H, Khanna H, Hildebrandt F: Candidate exome capture identifies mutation of SDCCAG8 as the cause of a retinal-renal ciliopathy. Nat Genet 42: 840–850, 2010 - PMC - PubMed
-
- Schaefer E, Zaloszyc A, Lauer J, Durand M, Stutzmann F, Perdomo-Trujillo Y, Redin C, Bennouna Greene V, Toutain A, Perrin L, Gérard M, Caillard S, Bei X, Lewis RA, Christmann D, Letsch J, Kribs M, Mutter C, Muller J, Stoetzel C, Fischbach M, Marion V, Katsanis N, Dollfus H: Mutations in SDCCAG8/NPHP10 cause Bardet-Biedl syndrome and are associated with penetrant renal disease and absent polydactyly. Mol Syndromol 1: 273–281, 2011 - PMC - PubMed
-
- Kenedy AA, Cohen KJ, Loveys DA, Kato GJ, Dang CV: Identification and characterization of the novel centrosome-associated protein CCCAP. Gene 303: 35–46, 2003 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
