

Microglial dynamics and role in the healthy and diseased brain: a paradigm of functional plasticity
- PMID: 24722525
- PMCID: PMC4412879
- DOI: 10.1177/1073858414530512
Microglial dynamics and role in the healthy and diseased brain: a paradigm of functional plasticity
Abstract
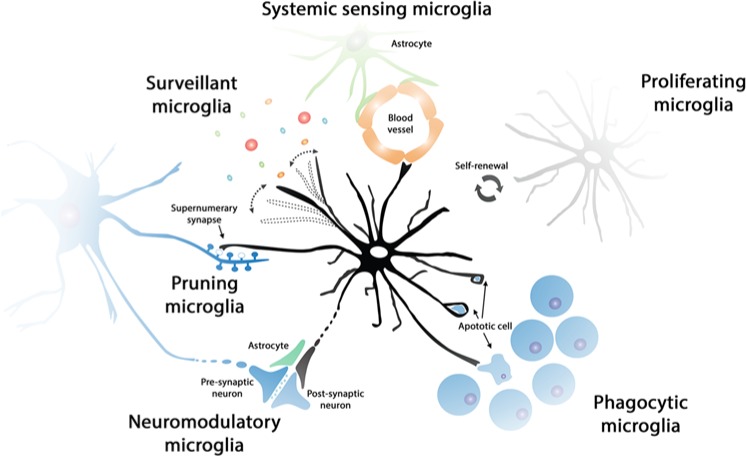
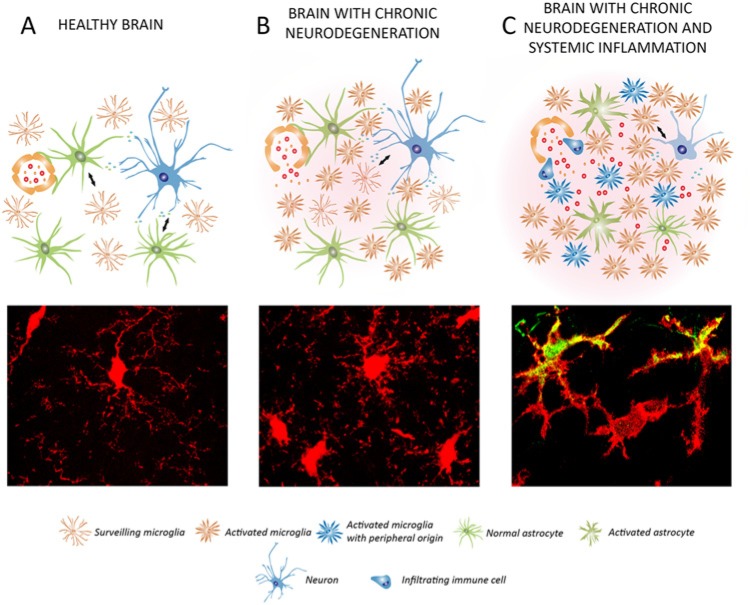
The study of the dynamics and functions of microglia in the healthy and diseased brain is a matter of intense scientific activity. The application of new techniques and new experimental approaches has allowed the identification of novel microglial functions and the redefinition of classic ones. In this review, we propose the study of microglial functions, rather than their molecular profiles, to better understand and define the roles of these cells in the brain. We review current knowledge on the role of surveillant microglia, proliferating microglia, pruning/neuromodulatory microglia, phagocytic microglia, and inflammatory microglia and the molecular profiles that are associated with these functions. In the remodeling scenario of microglial biology, the analysis of microglial functional states will inform about the roles in health and disease and will guide us to a more precise understanding of the multifaceted roles of this never-resting cells.
Keywords: microglia; neuroinflammation; neuromodulation; phagocytosis; proliferation; surveillance; synaptic pruning.
© The Author(s) 2014.
Conflict of interest statement
Figures

References
-
- Ajami B, Bennett JL, Krieger C, McNagny KM, Rossi FM. 2011. Infiltrating monocytes trigger EAE progression, but do not contribute to the resident microglia pool. Nat Neurosci 14(9):1142–9. - PubMed
-
- Ajami B, Bennett JL, Krieger C, Tetzlaff W, Rossi FM. 2007. Local self-renewal can sustain CNS microglia maintenance and function throughout adult life. Nat Neurosci 10(12):1538–43. - PubMed
-
- Akiyama H, Nishimura T, Kondo H, Ikeda K, Hayashi Y, McGeer PL. 1994. Expression of the receptor for macrophage colony stimulating factor by brain microglia and its upregulation in brains of patients with Alzheimer’s disease and amyotrophic lateral sclerosis. Brain Res 639(1):171–4. - PubMed
-
- Alliot F, Godin I, Pessac B. 1999. Microglia derive from progenitors, originating from the yolk sac, and which proliferate in the brain. Brain Res Dev Brain Res 117(2):145–52. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
