

Complement-related proteins control the flavivirus infection of Aedes aegypti by inducing antimicrobial peptides
- PMID: 24722701
- PMCID: PMC3983052
- DOI: 10.1371/journal.ppat.1004027
Complement-related proteins control the flavivirus infection of Aedes aegypti by inducing antimicrobial peptides
Abstract
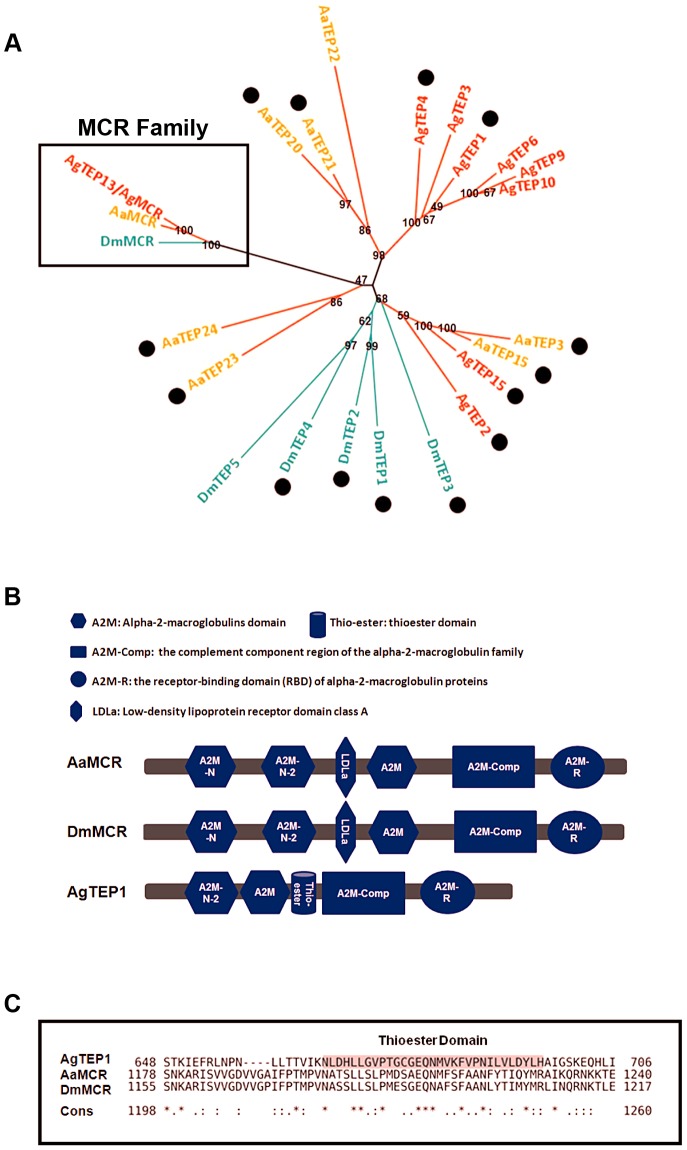
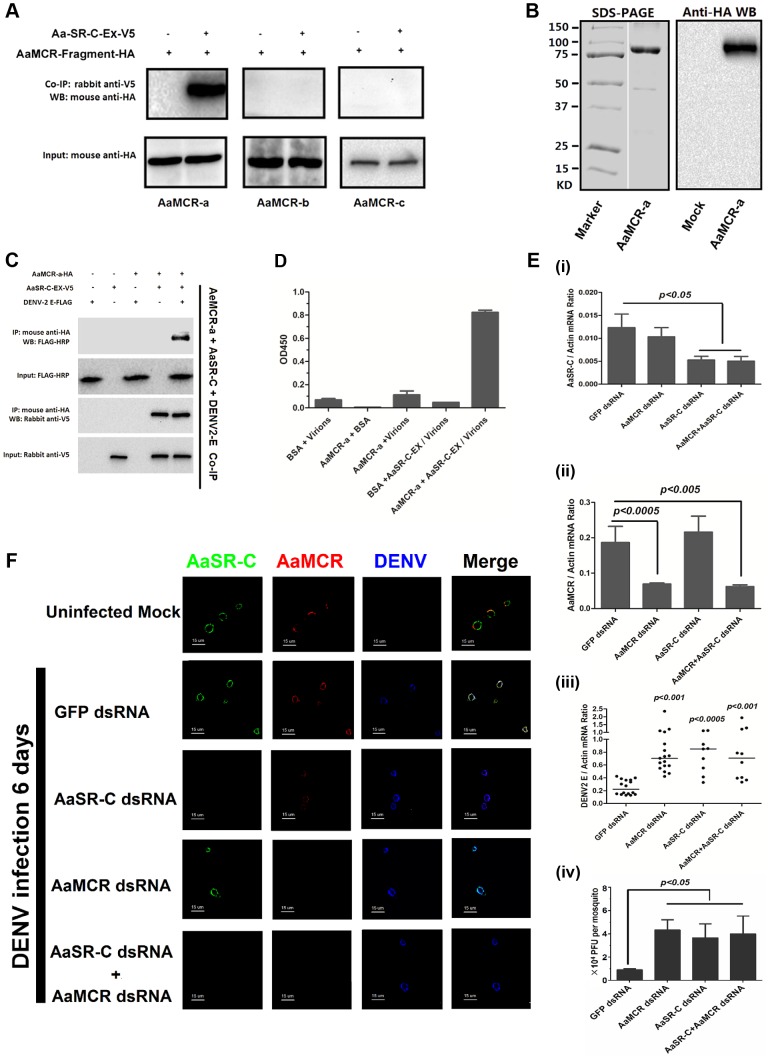
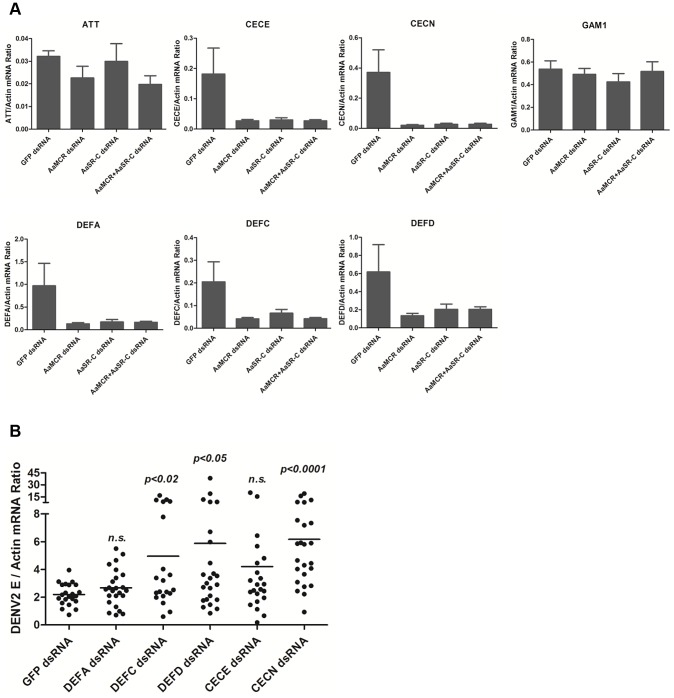
The complement system functions during the early phase of infection and directly mediates pathogen elimination. The recent identification of complement-like factors in arthropods indicates that this system shares common ancestry in vertebrates and invertebrates as an immune defense mechanism. Thioester (TE)-containing proteins (TEPs), which show high similarity to mammalian complement C3, are thought to play a key role in innate immunity in arthropods. Herein, we report that a viral recognition cascade composed of two complement-related proteins limits the flaviviral infection of Aedes aegypti. An A. aegypti macroglobulin complement-related factor (AaMCR), belonging to the insect TEP family, is a crucial effector in opposing the flaviviral infection of A. aegypti. However, AaMCR does not directly interact with DENV, and its antiviral effect requires an A. aegypti homologue of scavenger receptor-C (AaSR-C), which interacts with DENV and AaMCR simultaneously in vitro and in vivo. Furthermore, recognition of DENV by the AaSR-C/AaMCR axis regulates the expression of antimicrobial peptides (AMPs), which exerts potent anti-DENV activity. Our results both demonstrate the existence of a viral recognition pathway that controls the flaviviral infection by inducing AMPs and offer insights into a previously unappreciated antiviral function of the complement-like system in arthropods.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Levashina EA, Moita LF, Blandin S, Vriend G, Lagueux M, et al. (2001) Conserved role of a complement-like protein in phagocytosis revealed by dsRNA knockout in cultured cells of the mosquito, Anopheles gambiae . Cell 104: 709–718. - PubMed
-
- Blandin S, Shiao SH, Moita LF, Janse CJ, Waters AP, et al. (2004) Complement-like protein TEP1 is a determinant of vectorial capacity in the malaria vector Anopheles gambiae . Cell 116: 661–670. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
