

Lymphatic vessel insufficiency in hypercholesterolemic mice alters lipoprotein levels and promotes atherogenesis
- PMID: 24723556
- PMCID: PMC4031366
- DOI: 10.1161/ATVBAHA.114.302528
Lymphatic vessel insufficiency in hypercholesterolemic mice alters lipoprotein levels and promotes atherogenesis
Abstract
Objective: Lymphatic vessels collect extravasated fluid and proteins from tissues to blood circulation as well as play an essential role in lipid metabolism by taking up intestinal chylomicrons. Previous studies have shown that impairment of lymphatic vessel function causes lymphedema and fat accumulation, but clear connections between arterial pathologies and lymphatic vessels have not been described.
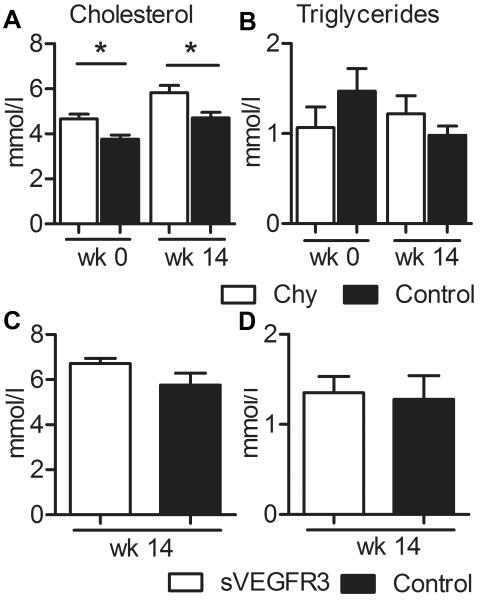
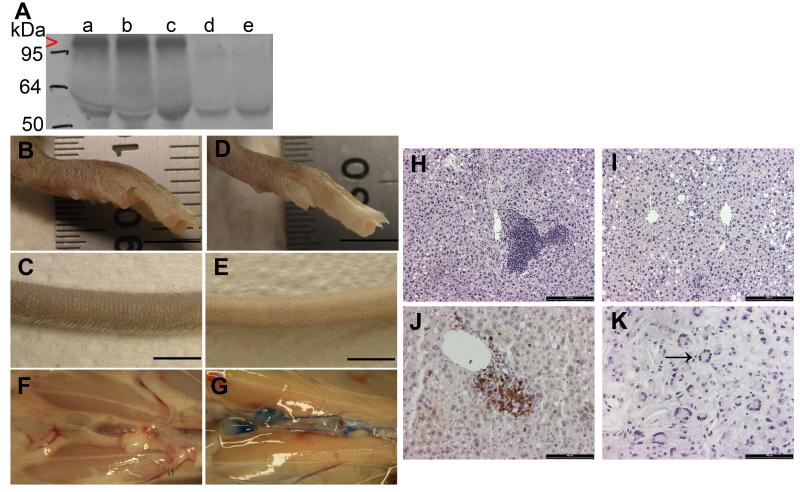
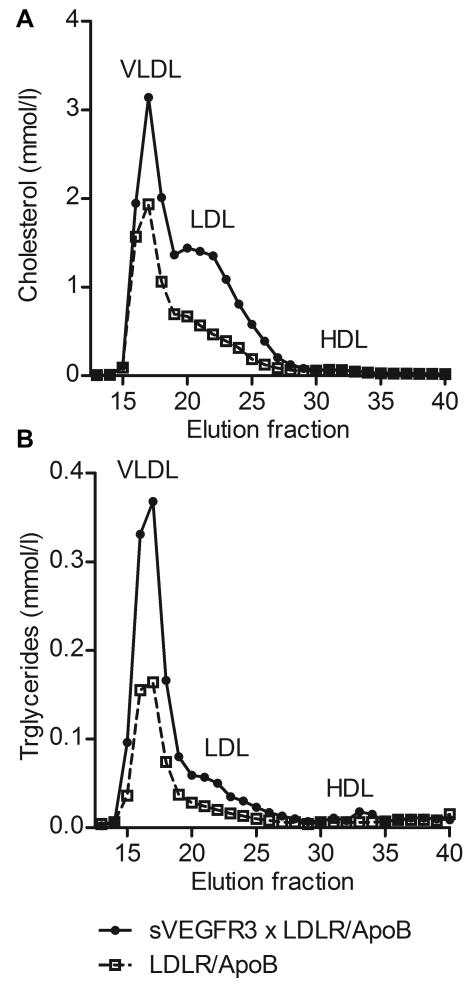
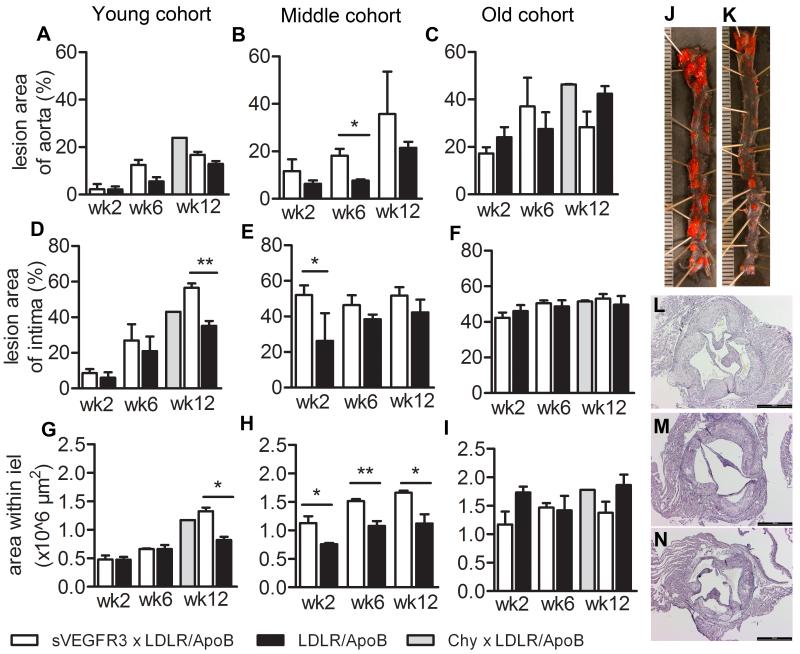
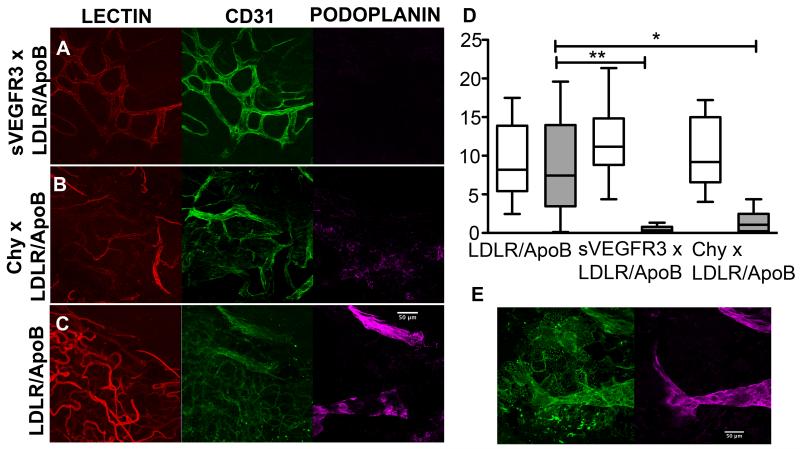
Approach and results: Two transgenic mouse strains with lymphatic insufficiency (soluble vascular endothelial growth factor 3 [sVEGFR3] and Chy) were crossed with atherosclerotic mice deficient of low-density lipoprotein receptor and apolipoprotein B48 (LDLR(-/-)/ApoB(100/100)) to study the effects of insufficient lymphatic vessel transport on lipoprotein metabolism and atherosclerosis. Both sVEGFR3×LDLR(-/-)/ApoB(100/100) mice and Chy×LDLR(-/-)/ApoB(100/100) mice had higher plasma cholesterol levels compared with LDLR(-/-)/ApoB(100/100) control mice during both normal chow diet (16.3 and 13.7 versus 8.2 mmol/L, respectively) and Western-type high-fat diet (eg, after 2 weeks of fat diet, 45.9 and 42.6 versus 30.2 mmol/L, respectively). Cholesterol and triglyceride levels in very-low-density lipoprotein and low-density lipoprotein fractions were increased. Atherosclerotic lesions in young and intermediate cohorts of sVEGFR3×LDLR(-/-)/ApoB(100/100) mice progressed faster than in control mice (eg, intermediate cohort mice at 6 weeks, 18.3% versus 7.7% of the whole aorta, respectively). In addition, lesions in sVEGFR3×LDLR(-/-)/ApoB(100/100) mice and Chy×LDLR(-/-)/ApoB(100/100) mice had much less lymphatic vessels than lesions in control mice (0.33% and 1.07% versus 7.45% of podoplanin-positive vessels, respectively).
Conclusions: We show a novel finding linking impaired lymphatic vessels to lipoprotein metabolism, increased plasma cholesterol levels, and enhanced atherogenesis.
Keywords: atherosclerosis; lipoproteins; lymphatic vessels.
© 2014 American Heart Association, Inc.
Figures

References
-
- Glass CK, Witztum JL. Atherosclerosis. the road ahead. Cell. 2001;104:503–516. - PubMed
-
- Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature. 1993;362:801–809. - PubMed
-
- Moulton KS. Angiogenesis in atherosclerosis: gathering evidence beyond speculation. Curr Opin Lipidol. 2006;17:548–555. - PubMed
-
- Yla-Herttuala S, Bentzon JF, Daemen M, et al. Stabilisation of atherosclerotic plaques. Position paper of the European Society of Cardiology (ESC) Working Group on atherosclerosis and vascular biology. Thromb Haemost. 2011;106:1–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
