

Atomistic autophagy: the structures of cellular self-digestion
- PMID: 24725401
- PMCID: PMC4038036
- DOI: 10.1016/j.cell.2014.01.070
Atomistic autophagy: the structures of cellular self-digestion
Abstract
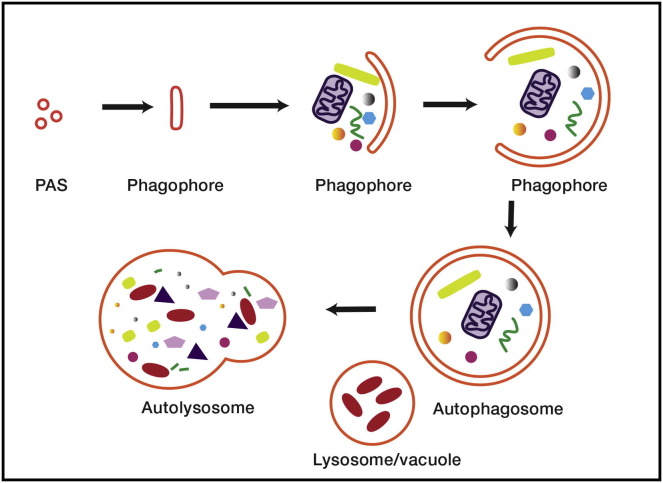
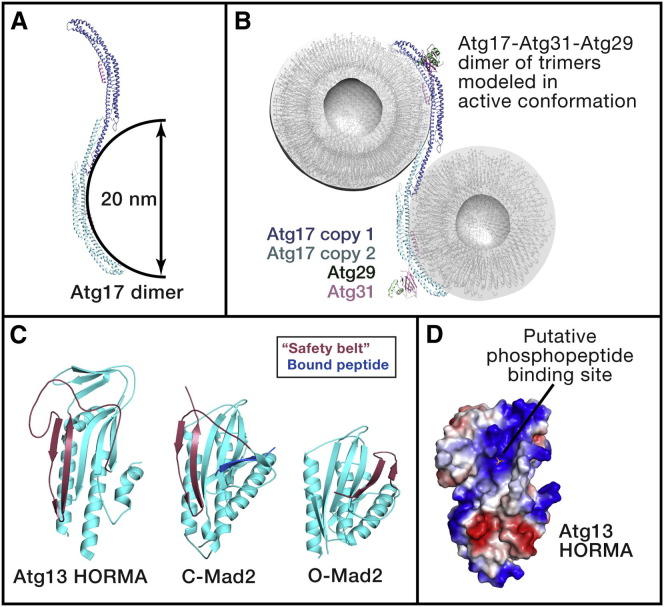
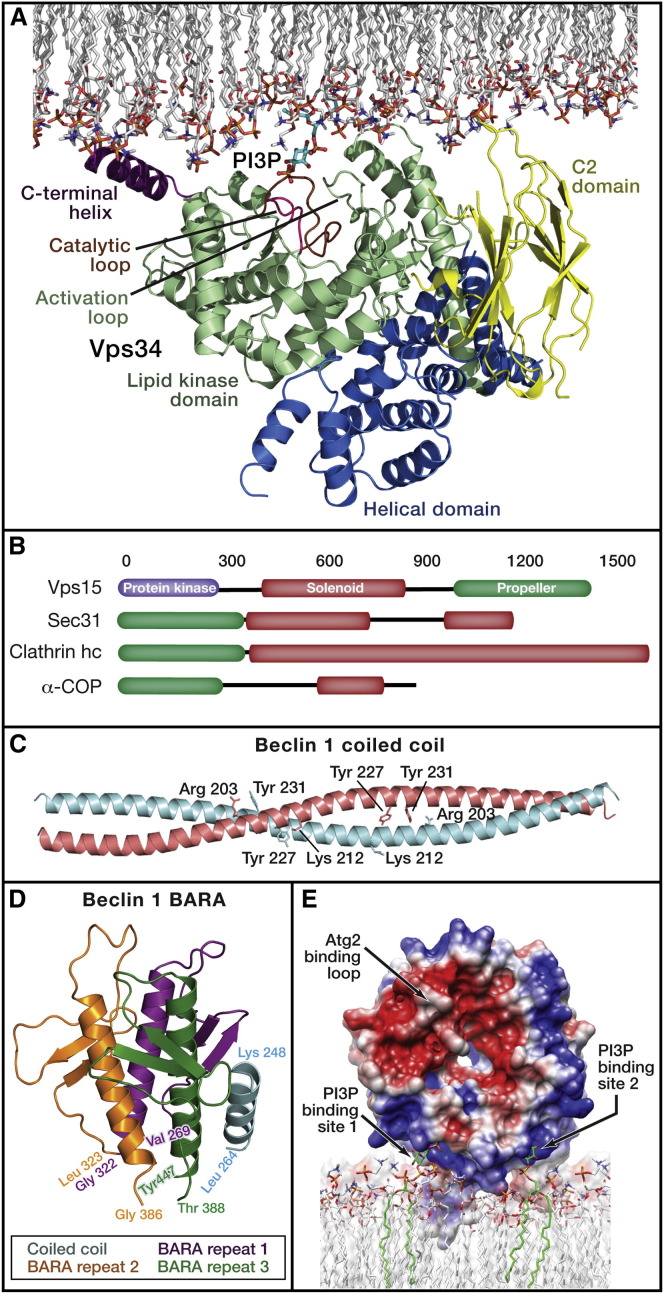
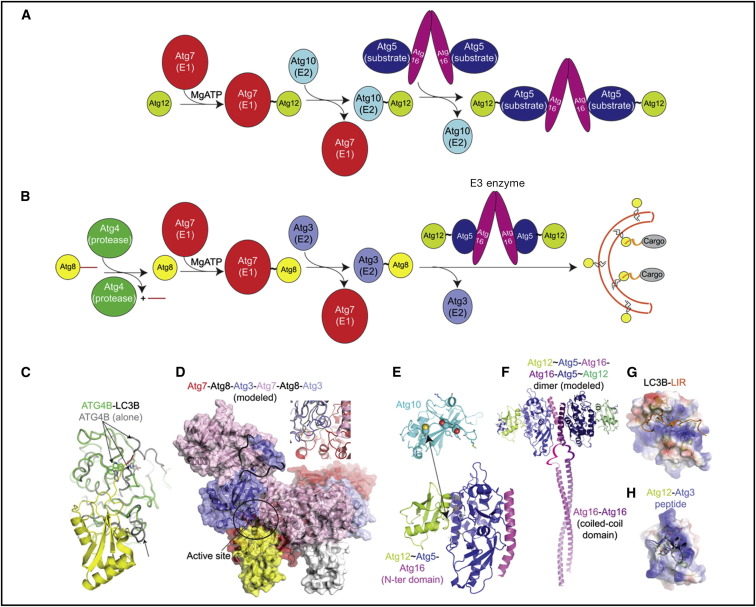
Autophagy is directed by numerous distinct autophagy-related (Atg) proteins. These transmit starvation-induced signals to lipids and regulatory proteins and assemble a double-membrane autophagosome sequestering bulk cytoplasm and/or selected cargos destined for degradation upon autophagosome fusion with a vacuole or lysosome. This Review discusses the structural mechanisms by which Atg proteins sense membrane curvature, mediate a PI(3)P-signaling cascade, and utilize autophagy-specific ubiquitin-like protein cascades to tether proteins to autophagosomal membranes. Recent elucidation of molecular interactions enabling vesicle nucleation, elongation, and cargo recruitment provides insights into how dynamic protein-protein and protein-membrane interactions may dictate size, shape, and contents of autophagosomes.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Mechanisms and regulation of autophagosome formation.Curr Opin Cell Biol. 2012 Aug;24(4):496-501. doi: 10.1016/j.ceb.2012.05.001. Epub 2012 Jun 2. Curr Opin Cell Biol. 2012. PMID: 22664348 Review.
-
Harpooning the Cvt complex to the phagophore assembly site.Autophagy. 2008 Oct;4(7):914-6. doi: 10.4161/auto.6657. Epub 2008 Oct 24. Autophagy. 2008. PMID: 18708760
-
The Atg17-Atg31-Atg29 complex and Atg11 regulate autophagosome-vacuole fusion.Autophagy. 2016 May 3;12(5):894-5. doi: 10.1080/15548627.2016.1162364. Autophagy. 2016. PMID: 26986547 Free PMC article. Review.
-
A current perspective of autophagosome biogenesis.Cell Res. 2014 Jan;24(1):58-68. doi: 10.1038/cr.2013.159. Epub 2013 Dec 3. Cell Res. 2014. PMID: 24296784 Free PMC article. Review.
-
Membrane Trafficking in Autophagy.Int Rev Cell Mol Biol. 2018;336:1-92. doi: 10.1016/bs.ircmb.2017.07.001. Epub 2017 Sep 21. Int Rev Cell Mol Biol. 2018. PMID: 29413888 Review.
Cited by
-
The Lysosome as a Regulatory Hub.Annu Rev Cell Dev Biol. 2016 Oct 6;32:223-253. doi: 10.1146/annurev-cellbio-111315-125125. Epub 2016 Aug 3. Annu Rev Cell Dev Biol. 2016. PMID: 27501449 Free PMC article. Review.
-
Regulation of LC3 lipidation by the autophagy-specific class III phosphatidylinositol-3 kinase complex.Mol Biol Cell. 2019 Apr 15;30(9):1098-1107. doi: 10.1091/mbc.E18-11-0743. Epub 2019 Feb 27. Mol Biol Cell. 2019. PMID: 30811270 Free PMC article.
-
Endoplasmic reticulum stress-mediated autophagy protects against β,β-dimethylacrylshikonin-induced apoptosis in lung adenocarcinoma cells.Cancer Sci. 2018 Jun;109(6):1889-1901. doi: 10.1111/cas.13616. Epub 2018 May 19. Cancer Sci. 2018. PMID: 29676829 Free PMC article.
-
[Changes in autophagy during maturation and differentiation of Hepa1-6 cells induced by all-trans retinoic acid].Nan Fang Yi Ke Da Xue Xue Bao. 2018 May 20;38(5):527-533. doi: 10.3969/j.issn.1673-4254.2018.05.04. Nan Fang Yi Ke Da Xue Xue Bao. 2018. PMID: 29891447 Free PMC article. Chinese.
-
A New Long Noncoding RNA ALB Regulates Autophagy by Enhancing the Transformation of LC3BI to LC3BII during Human Lens Development.Mol Ther Nucleic Acids. 2017 Dec 15;9:207-217. doi: 10.1016/j.omtn.2017.09.011. Epub 2017 Oct 5. Mol Ther Nucleic Acids. 2017. PMID: 29246299 Free PMC article.
References
-
- Brownell J.E., Sintchak M.D., Gavin J.M., Liao H., Bruzzese F.J., Bump N.J., Soucy T.A., Milhollen M.A., Yang X., Burkhardt A.L. Substrate-assisted inhibition of ubiquitin-like protein-activating enzymes: the NEDD8 E1 inhibitor MLN4924 forms a NEDD8-AMP mimetic in situ. Mol. Cell. 2010;37:102–111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
