

Intestinal brush border assembly driven by protocadherin-based intermicrovillar adhesion
- PMID: 24725409
- PMCID: PMC3992856
- DOI: 10.1016/j.cell.2014.01.067
Intestinal brush border assembly driven by protocadherin-based intermicrovillar adhesion
Abstract
Transporting epithelial cells build apical microvilli to increase membrane surface area and enhance absorptive capacity. The intestinal brush border provides an elaborate example with tightly packed microvilli that function in nutrient absorption and host defense. Although the brush border is essential for physiological homeostasis, its assembly is poorly understood. We found that brush border assembly is driven by the formation of Ca(2+)-dependent adhesion links between adjacent microvilli. Intermicrovillar links are composed of protocadherin-24 and mucin-like protocadherin, which target to microvillar tips and interact to form a trans-heterophilic complex. The cytoplasmic domains of microvillar protocadherins interact with the scaffolding protein, harmonin, and myosin-7b, which promote localization to microvillar tips. Finally, a mouse model of Usher syndrome lacking harmonin exhibits microvillar protocadherin mislocalization and severe defects in brush border morphology. These data reveal an adhesion-based mechanism for brush border assembly and illuminate the basis of intestinal pathology in patients with Usher syndrome. PAPERFLICK:
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
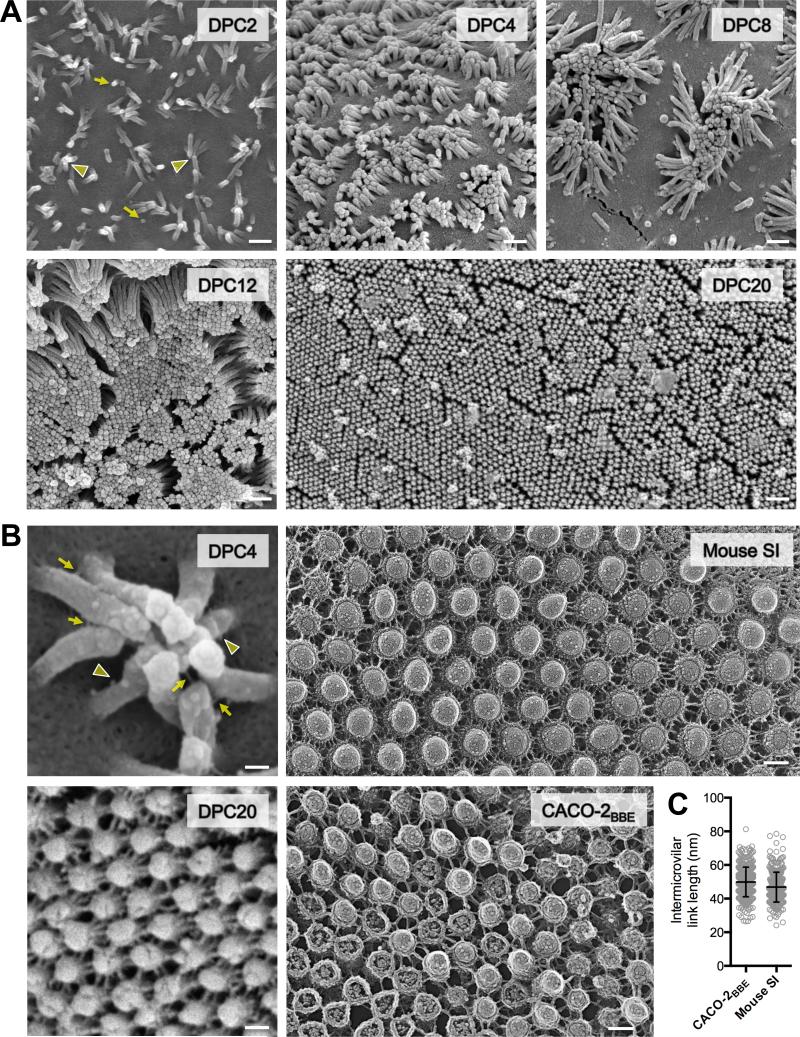
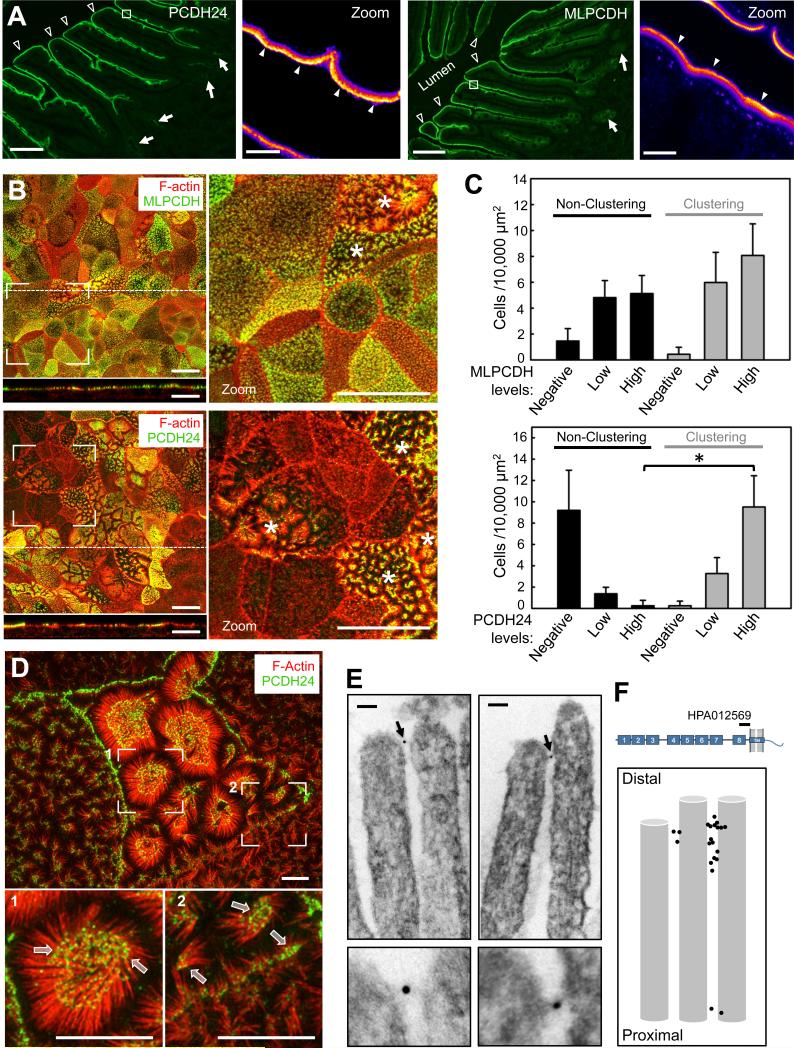
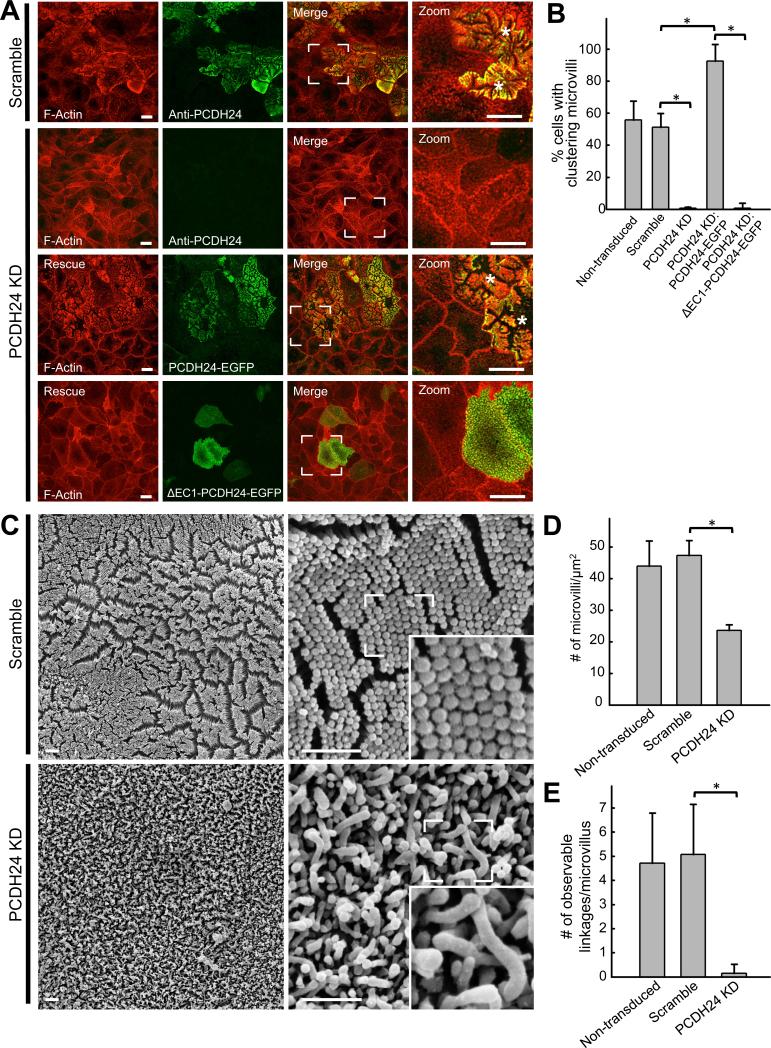
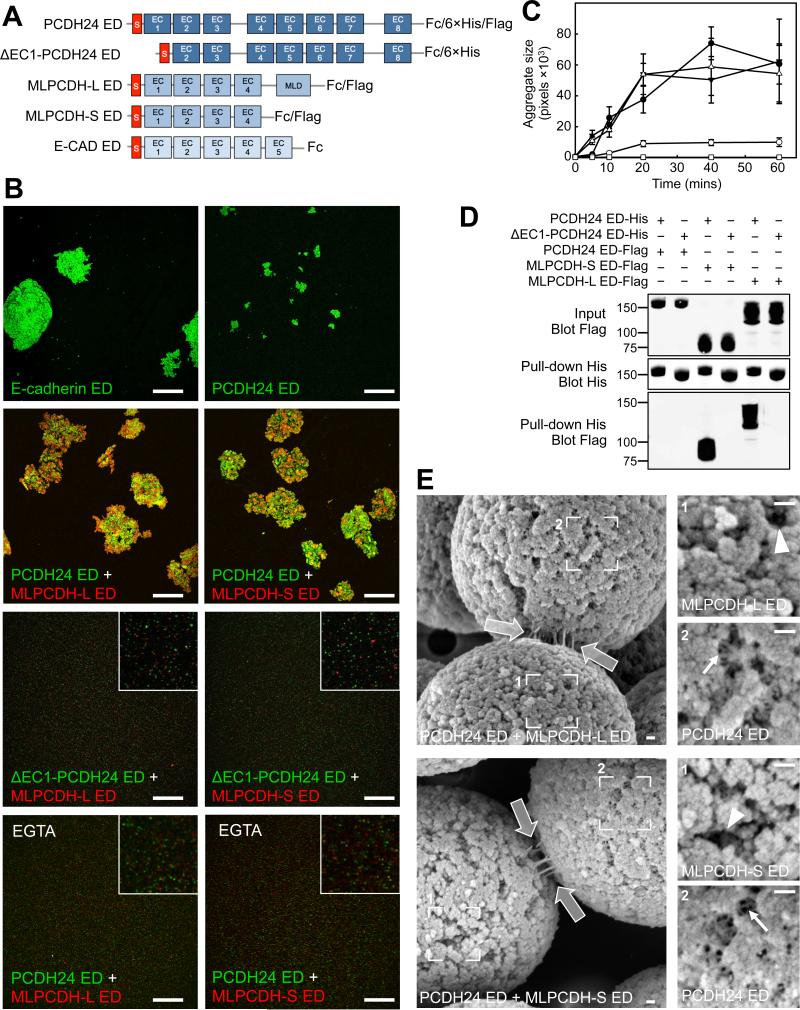
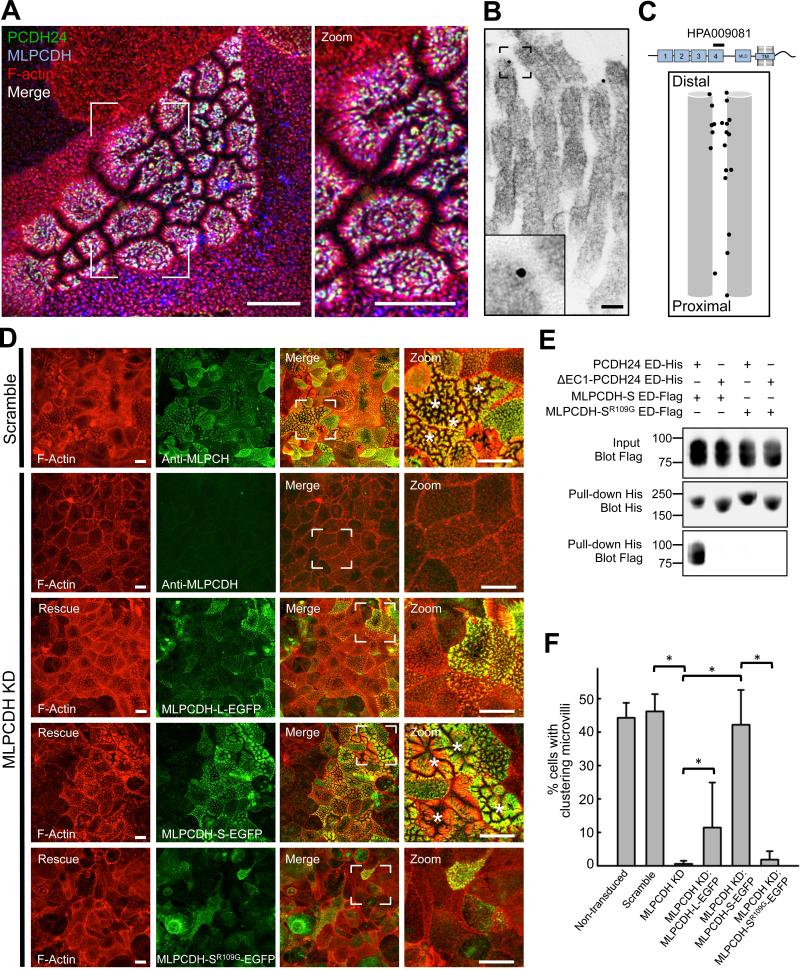
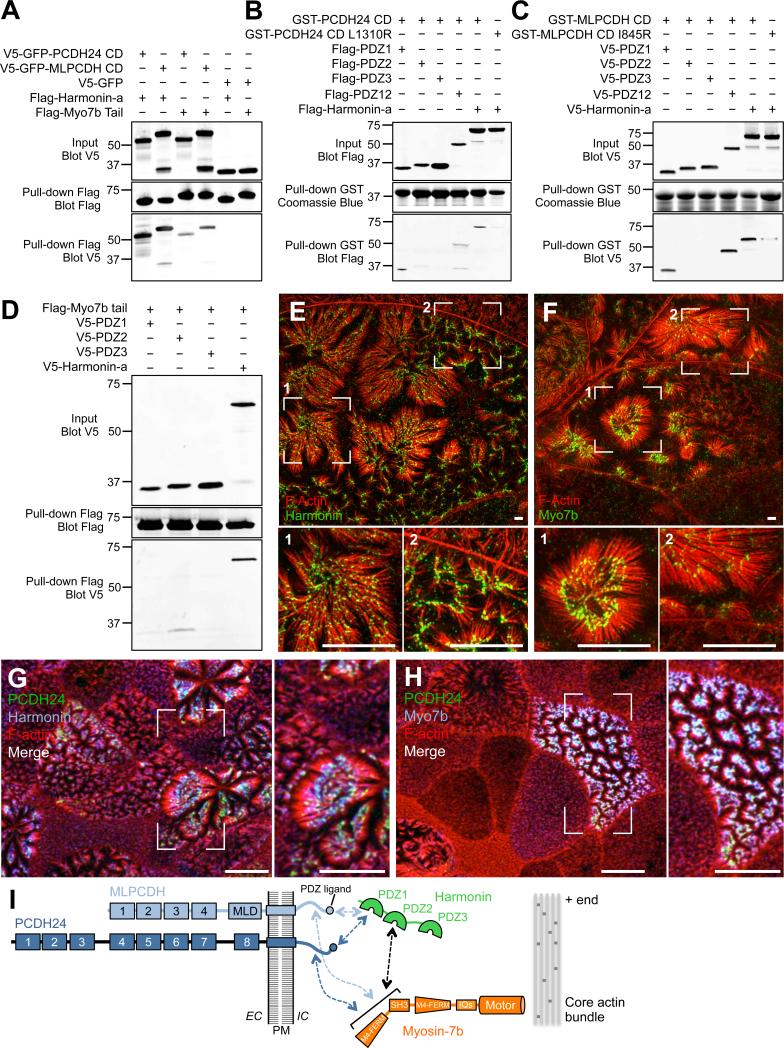
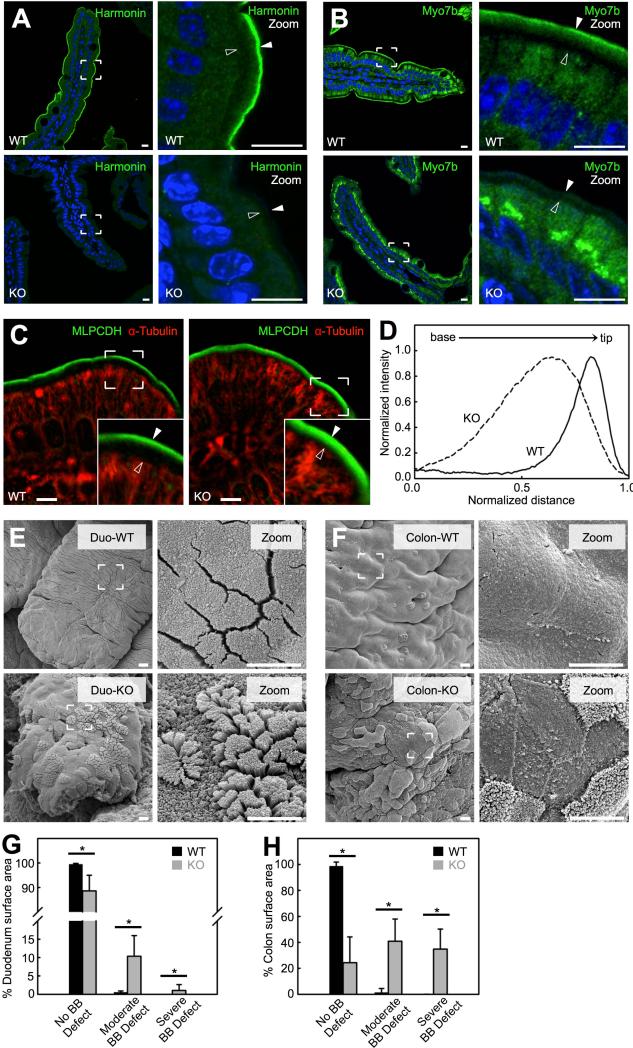
References
-
- Belyantseva IA, Boger ET, Naz S, Frolenkov GI, Sellers JR, Ahmed ZM, Griffith AJ, Friedman TB. Myosin-XVa is required for tip localization of whirlin and differential elongation of hair-cell stereocilia. Nat Cell Biol. 2005;7:148–156. - PubMed
-
- Bitner-Glindzicz M, Lindley KJ, Rutland P, Blaydon D, Smith VV, Milla PJ, Hussain K, Furth-Lavi J, Cosgrove KE, Shepherd RM, et al. A recessive contiguous gene deletion causing infantile hyperinsulinism, enteropathy and deafness identifies the Usher type 1C gene. Nat Genet. 2000;26:56–60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R56 DK095811/DK/NIDDK NIH HHS/United States
- T32 HD007502/HD/NICHD NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- DK095811/DK/NIDDK NIH HHS/United States
- R01 DC009246/DC/NIDCD NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- T32 GM008554/GM/NIGMS NIH HHS/United States
- P30DK058404/DK/NIDDK NIH HHS/United States
- DK075555/DK/NIDDK NIH HHS/United States
- S10 OD012324/OD/NIH HHS/United States
- T32 CA119925/CA/NCI NIH HHS/United States
- R01 DK095811/DK/NIDDK NIH HHS/United States
- R01 DK075555/DK/NIDDK NIH HHS/United States
- DC009246/DC/NIDCD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
