

Reprogramming acyl carrier protein interactions of an Acyl-CoA promiscuous trans-acyltransferase
- PMID: 24726832
- PMCID: PMC4035391
- DOI: 10.1016/j.chembiol.2014.02.019
Reprogramming acyl carrier protein interactions of an Acyl-CoA promiscuous trans-acyltransferase
Abstract
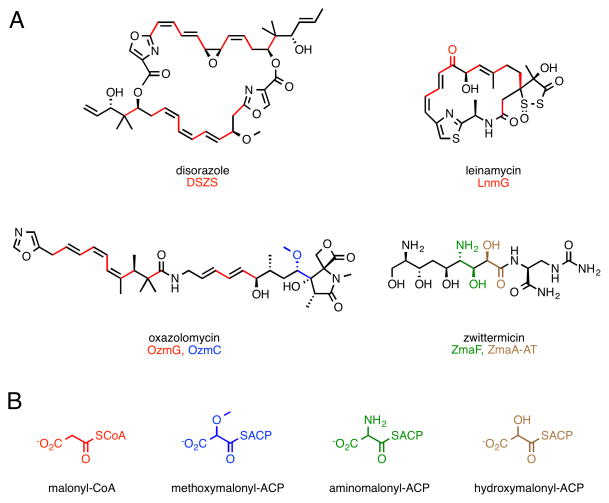
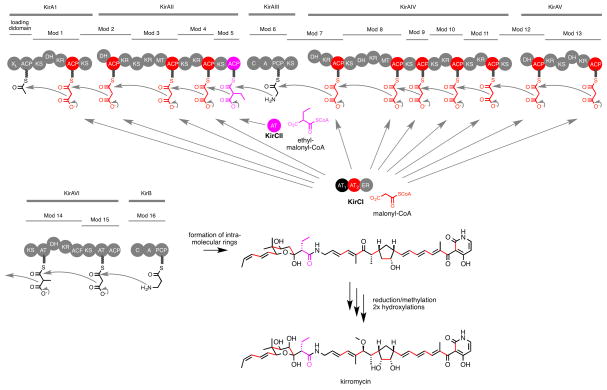
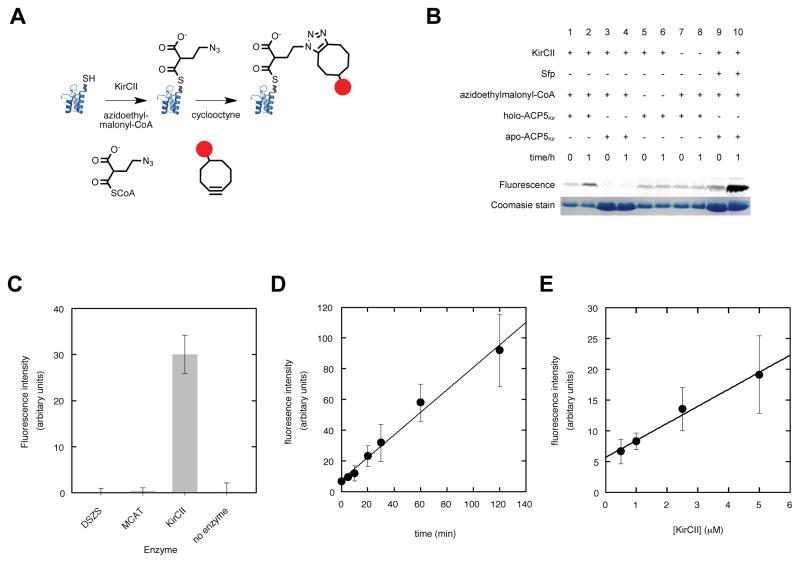
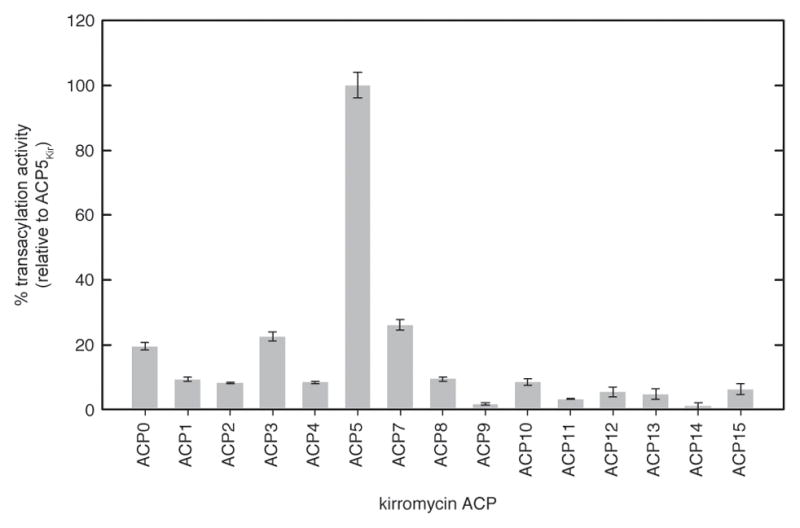
Protein interactions between acyl carrier proteins (ACPs) and trans-acting acyltransferase domains (trans-ATs) are critical for regioselective extender unit installation by many polyketide synthases, yet little is known regarding the specificity of these interactions, particularly for trans-ATs with unusual extender unit specificities. Currently, the best-studied trans-AT with nonmalonyl specificity is KirCII from kirromycin biosynthesis. Here, we developed an assay to probe ACP interactions based on leveraging the extender unit promiscuity of KirCII. The assay allows us to identify residues on the ACP surface that contribute to specific recognition by KirCII. This information proved sufficient to modify a noncognate ACP from a different biosynthetic system to be a substrate for KirCII. The findings form a foundation for further understanding the specificity of trans-AT:ACP protein interactions and for engineering modular polyketide synthases to produce analogs.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures

Similar articles
-
Supramolecular templating in kirromycin biosynthesis: the acyltransferase KirCII loads ethylmalonyl-CoA extender onto a specific ACP of the trans-AT PKS.Chem Biol. 2011 Apr 22;18(4):438-44. doi: 10.1016/j.chembiol.2011.02.007. Chem Biol. 2011. PMID: 21513880
-
Comparative analysis of the substrate specificity of trans- versus cis-acyltransferases of assembly line polyketide synthases.Biochemistry. 2014 Jun 17;53(23):3796-806. doi: 10.1021/bi5004316. Epub 2014 Jun 9. Biochemistry. 2014. PMID: 24871074 Free PMC article.
-
Extender Unit Promiscuity and Orthogonal Protein Interactions of an Aminomalonyl-ACP Utilizing Trans-Acyltransferase from Zwittermicin Biosynthesis.ACS Chem Biol. 2018 Dec 21;13(12):3361-3373. doi: 10.1021/acschembio.8b00867. Epub 2018 Nov 28. ACS Chem Biol. 2018. PMID: 30484625 Free PMC article.
-
Acyl-coenzyme A:(holo-acyl carrier protein) transacylase enzymes as templates for engineering.Appl Microbiol Biotechnol. 2018 Aug;102(15):6333-6341. doi: 10.1007/s00253-018-9114-2. Epub 2018 Jun 1. Appl Microbiol Biotechnol. 2018. PMID: 29858956 Review.
-
Probing the structure and function of acyl carrier proteins to unlock the strategic redesign of type II polyketide biosynthetic pathways.J Biol Chem. 2021 Jan-Jun;296:100328. doi: 10.1016/j.jbc.2021.100328. Epub 2021 Jan 23. J Biol Chem. 2021. PMID: 33493513 Free PMC article. Review.
Cited by
-
A dual transacylation mechanism for polyketide synthase chain release in enacyloxin antibiotic biosynthesis.Nat Chem. 2019 Oct;11(10):906-912. doi: 10.1038/s41557-019-0309-7. Epub 2019 Sep 23. Nat Chem. 2019. PMID: 31548673 Free PMC article.
-
Branched late-steps of the cytosolic iron-sulphur cluster assembly machinery of Trypanosoma brucei.PLoS Pathog. 2018 Oct 22;14(10):e1007326. doi: 10.1371/journal.ppat.1007326. eCollection 2018 Oct. PLoS Pathog. 2018. PMID: 30346997 Free PMC article.
-
Harnessing natural product assembly lines: structure, promiscuity, and engineering.J Ind Microbiol Biotechnol. 2016 Mar;43(2-3):371-87. doi: 10.1007/s10295-015-1704-8. Epub 2015 Nov 2. J Ind Microbiol Biotechnol. 2016. PMID: 26527577 Free PMC article. Review.
-
Acyltransferases as Tools for Polyketide Synthase Engineering.Antibiotics (Basel). 2018 Jul 18;7(3):62. doi: 10.3390/antibiotics7030062. Antibiotics (Basel). 2018. PMID: 30022008 Free PMC article. Review.
-
Engineered polyketides: Synergy between protein and host level engineering.Synth Syst Biotechnol. 2017 Sep 7;2(3):147-166. doi: 10.1016/j.synbio.2017.08.005. eCollection 2017 Sep. Synth Syst Biotechnol. 2017. PMID: 29318196 Free PMC article. Review.
References
-
- Arnold K, Bordoli L, Kopp J, Schwede T. The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics. 2006;22:195–201. - PubMed
-
- Arthur CJ, Williams C, Pottage K, Ploskon E, Findlow SC, Burston SG, Simpson TJ, Crump MP, Crosby J. Structure and malonyl CoA-ACP transacylase binding of streptomyces coelicolor fatty acid synthase acyl carrier protein. ACS Chem Biol. 2009;4:625–636. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
