

Alterations in cerebellar physiology are associated with a stiff-legged gait in Atcay(ji-hes) mice
- PMID: 24727095
- PMCID: PMC4059535
- DOI: 10.1016/j.nbd.2014.03.020
Alterations in cerebellar physiology are associated with a stiff-legged gait in Atcay(ji-hes) mice
Abstract
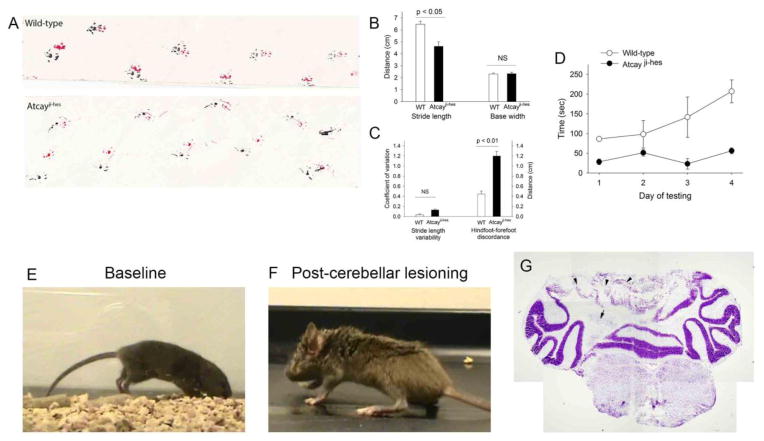
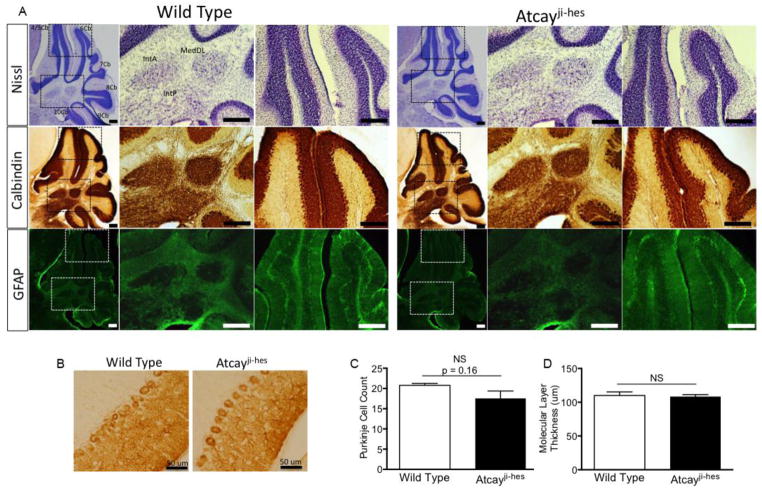
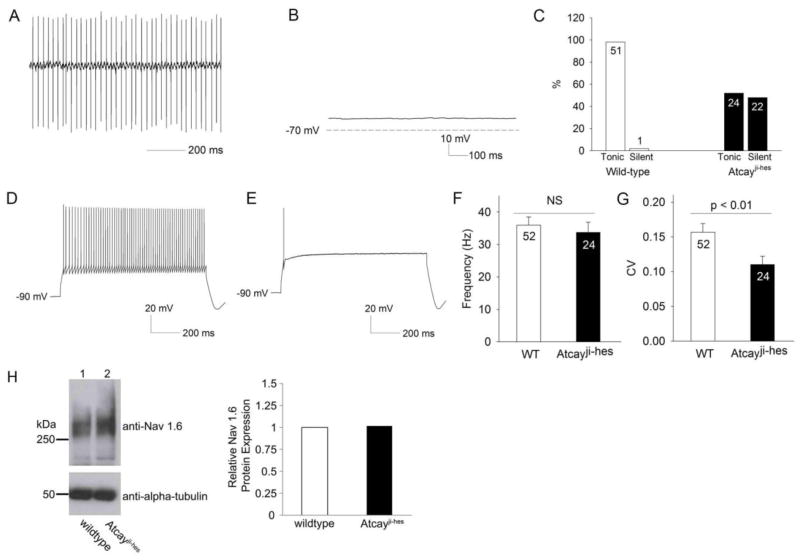
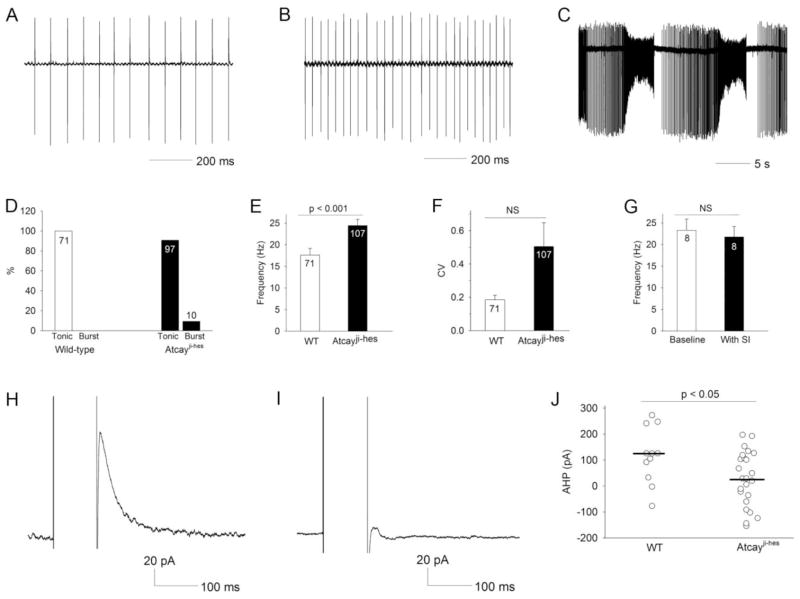
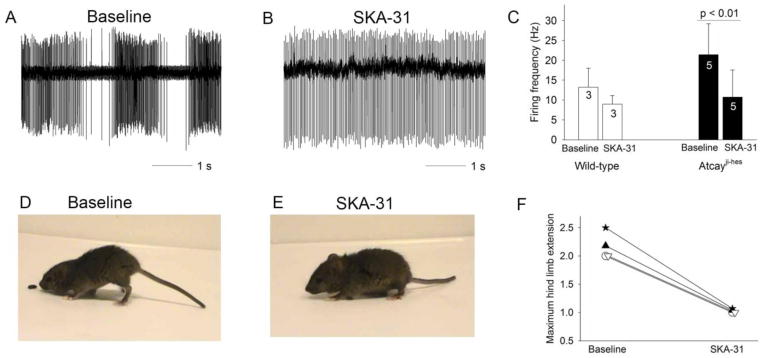
Recent evidence suggests that dystonia, a movement disorder characterized by sustained involuntary muscle contractions, can be associated with cerebellar abnormalities. The basis for how functional changes in the cerebellum can cause dystonia is poorly understood. Here we identify alterations in physiology in Atcay(ji-hes) mice which in addition to ataxia, have an abnormal gait with hind limb extension and toe walking, reminiscent of human dystonic gait. No morphological abnormalities in the brain accompany the dystonia, but partial cerebellectomy causes resolution of the stiff-legged gait, suggesting that cerebellar dysfunction contributes to the dystonic gait of Atcay(ji-hes) mice. Recordings from Purkinje and deep cerebellar nuclear (DCN) neurons in acute brain slices were used to determine the physiological correlates of dystonia in the Atcay(ji-hes) mice. Approximately 50% of cerebellar Purkinje neurons fail to display the normal repetitive firing characteristic of these cells. In addition, DCN neurons exhibit increased intrinsic firing frequencies with a subset of neurons displaying bursts of action potentials. This increased intrinsic excitability of DCN neurons is accompanied by a reduction in after-hyperpolarization currents mediated by small-conductance calcium-activated potassium (SK) channels. An activator of SK channels reduces DCN neuron firing frequency in acute cerebellar slices and improves the dystonic gait of Atcay(ji-hes) mice. These results suggest that a combination of reduced Purkinje neuron activity and increased DCN intrinsic excitability can result in a combination of ataxia and a dystonia-like gait in mice.
Keywords: Ataxia; Cerebellum; Deep cerebellar nuclei; Dystonia; Electrophysiology; Mutant mice; Patch-clamp; Purkinje cells.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Unusually Slow Spike Frequency Adaptation in Deep Cerebellar Nuclei Neurons Preserves Linear Transformations on the Subsecond Timescale.J Neurosci. 2022 Oct 5;42(40):7581-7593. doi: 10.1523/JNEUROSCI.1869-21.2022. Epub 2022 Aug 22. J Neurosci. 2022. PMID: 35995561 Free PMC article.
-
Enhanced neuronal excitability in the absence of neurodegeneration induces cerebellar ataxia.J Clin Invest. 2004 Feb;113(4):582-90. doi: 10.1172/JCI20216. J Clin Invest. 2004. PMID: 14966567 Free PMC article.
-
Abnormal high-frequency burst firing of cerebellar neurons in rapid-onset dystonia-parkinsonism.J Neurosci. 2014 Aug 27;34(35):11723-32. doi: 10.1523/JNEUROSCI.1409-14.2014. J Neurosci. 2014. PMID: 25164667 Free PMC article.
-
Increased Purkinje Cell Complex Spike and Deep Cerebellar Nucleus Synchrony as a Potential Basis for Syndromic Essential Tremor. A Review and Synthesis of the Literature.Cerebellum. 2021 Apr;20(2):266-281. doi: 10.1007/s12311-020-01197-5. Epub 2020 Oct 13. Cerebellum. 2021. PMID: 33048308 Review.
-
Population coding in the cerebellum: a machine learning perspective.J Neurophysiol. 2020 Dec 1;124(6):2022-2051. doi: 10.1152/jn.00449.2020. Epub 2020 Oct 28. J Neurophysiol. 2020. PMID: 33112717 Free PMC article. Review.
Cited by
-
Persistent motor dysfunction despite homeostatic rescue of cerebellar morphogenesis in the Car8 waddles mutant mouse.Neural Dev. 2019 Mar 12;14(1):6. doi: 10.1186/s13064-019-0130-4. Neural Dev. 2019. PMID: 30867000 Free PMC article.
-
Dissecting the links between cerebellum and dystonia.Cerebellum. 2014 Dec;13(6):666-8. doi: 10.1007/s12311-014-0601-4. Cerebellum. 2014. PMID: 25239288
-
A Case of Masticatory Dystonia Following Cerebellar Haemorrhage.Cerebellum. 2015 Dec;14(6):723-7. doi: 10.1007/s12311-015-0655-y. Cerebellum. 2015. PMID: 25700680 No abstract available.
-
Structural insights into the potency of SK channel positive modulators.Sci Rep. 2017 Dec 7;7(1):17178. doi: 10.1038/s41598-017-16607-8. Sci Rep. 2017. PMID: 29214998 Free PMC article.
-
Abnormal cerebellar function and tremor in a mouse model for non-manifesting partially penetrant dystonia type 6.J Physiol. 2021 Apr;599(7):2037-2054. doi: 10.1113/JP280978. Epub 2021 Jan 9. J Physiol. 2021. PMID: 33369735 Free PMC article.
References
-
- Aizenman CD, Linden DJ. Regulation of the rebound depolarization and spontaneous firing patterns of deep nuclear neurons in slices of rat cerebellum. J Neurophysiol. 1999;82:1697–709. - PubMed
-
- Alvarez-Fischer D, et al. Prolonged generalized dystonia after chronic cerebellar application of kainic acid. Brain Res. 2012;1464:82–8. - PubMed
-
- Arif B, et al. An unusual neurological syndrome of crawling gait, dystonia, pyramidal signs, and limited speech. Mov Disord. 2011;26:2279–83. - PubMed
-
- Berardelli A, et al. The pathophysiology of primary dystonia. Brain. 1998;121(Pt 7):1195–212. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
