

Hc-daf-2 encodes an insulin-like receptor kinase in the barber's pole worm, Haemonchus contortus, and restores partial dauer regulation
- PMID: 24727120
- PMCID: PMC4516220
- DOI: 10.1016/j.ijpara.2014.03.005
Hc-daf-2 encodes an insulin-like receptor kinase in the barber's pole worm, Haemonchus contortus, and restores partial dauer regulation
Abstract
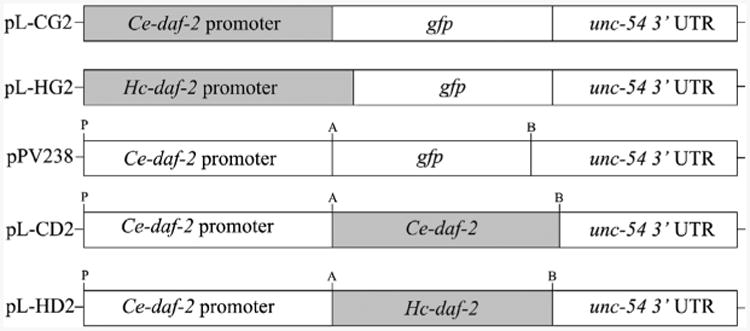
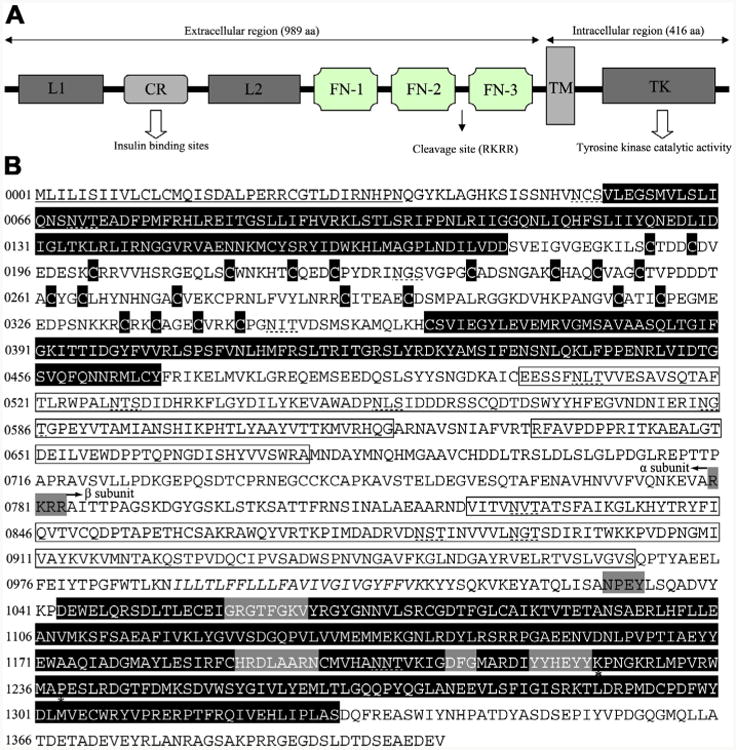
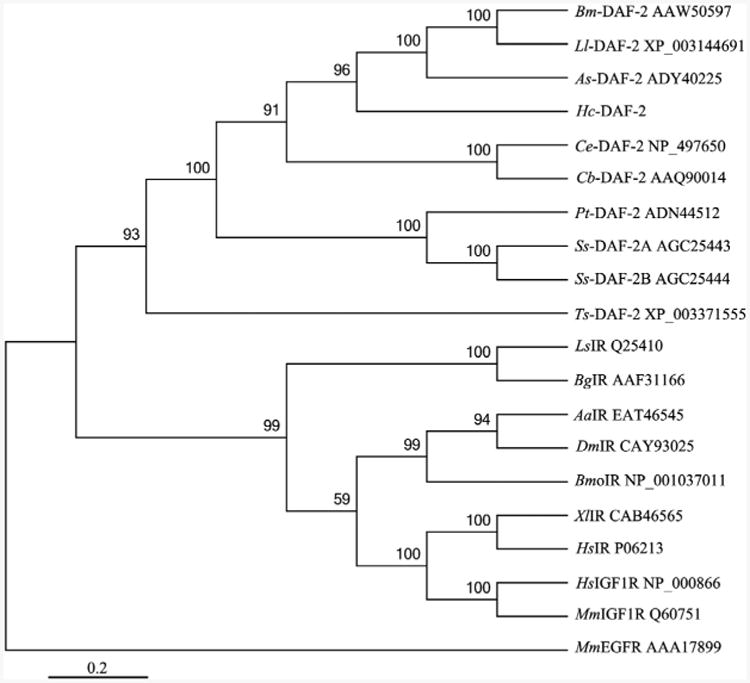
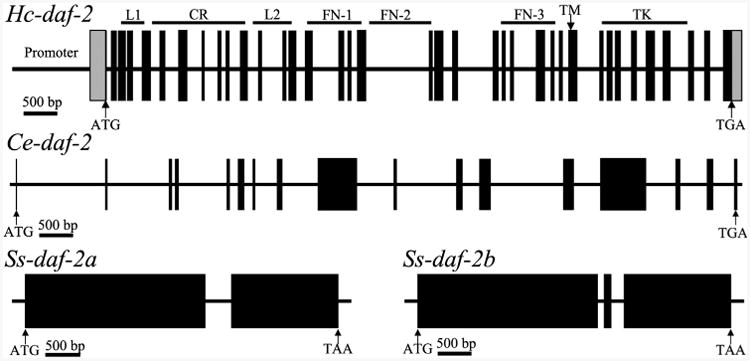
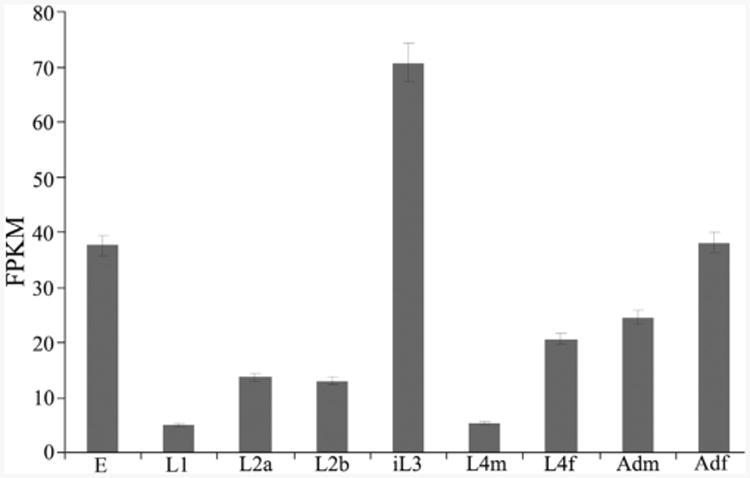
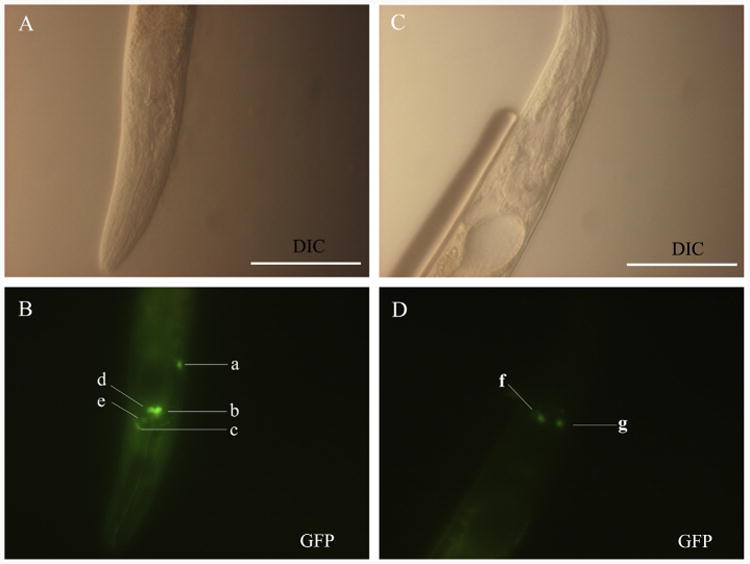
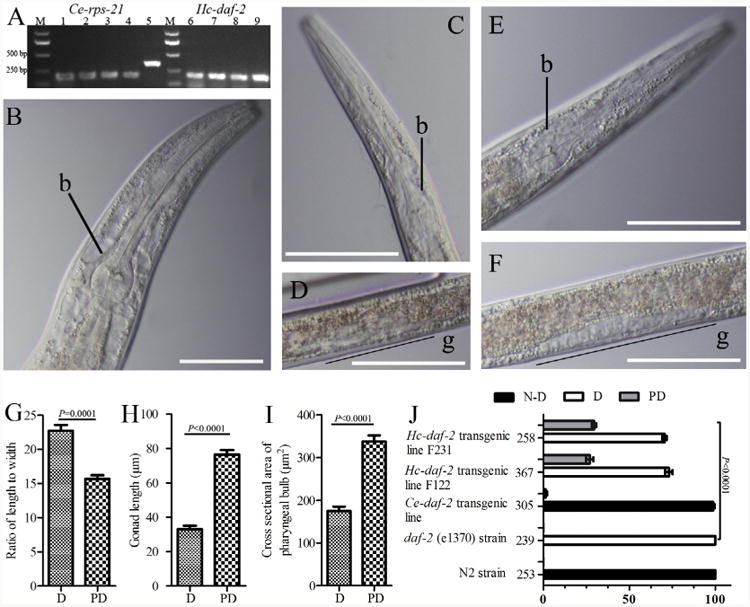
Infective L3s (iL3s) of parasitic nematodes share common behavioural, morphological and developmental characteristics with the developmentally arrested (dauer) larvae of the free-living nematode Caenorhabditis elegans. It is proposed that similar molecular mechanisms regulate entry into or exit from the dauer stage in C. elegans, and the transition from free-living to parasitic forms of parasitic nematodes. In C. elegans, one of the key factors regulating the dauer transition is the insulin-like receptor (designated Ce-DAF-2) encoded by the gene Ce-daf-2. However, nothing is known about DAF-2 homologues in most parasitic nematodes. Here, using a PCR-based approach, we identified and characterised a gene (Hc-daf-2) and its inferred product (Hc-DAF-2) in Haemonchus contortus (a socioeconomically important parasitic nematode of ruminants). The sequence of Hc-DAF-2 displays significant sequence homology to insulin receptors (IR) in both vertebrates and invertebrates, and contains conserved structural domains. A sequence encoding an important proteolytic motif (RKRR) identified in the predicted peptide sequence of Hc-DAF-2 is consistent with that of the human IR, suggesting that it is involved in the formation of the IR complex. The Hc-daf-2 gene was transcribed in all life stages of H. contortus, with a significant up-regulation in the iL3 compared with other stages. To compare patterns of expression between Hc-daf-2 and Ce-daf-2, reporter constructs fusing the Ce-daf-2 or Hc-daf-2 promoter to sequence encoding GFP were microinjected into the N2 strain of C. elegans, and transgenic lines were established and examined. Both genes showed similar patterns of expression in amphidial (head) neurons, which relate to sensation and signal transduction. Further study by heterologous genetic complementation in a daf-2-deficient strain of C. elegans (CB1370) showed partial rescue of function by Hc-daf-2. Taken together, these findings provide a first insight into the roles of Hc-daf-2/Hc-DAF-2 in the biology and development of H. contortus, particularly in the transition to parasitism.
Keywords: Development; Haemonchus contortus; Parasitic nematode; Transgenesis; daf-2.
Copyright © 2014 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Figures
References
-
- Aboobaker AA, Blaxter ML. Functional genomics for parasitic nematodes and platyhelminths. Trends Parasitol. 2004;20:178–184. - PubMed
-
- Apfeld J, Kenyon C. Cell nonautonomy of C. elegans daf-2 function in the regulation of diapause and life span. Cell. 1998;95:199–210. - PubMed
-
- Bargmann CI. Chemosensation in C. elegans. WormBook, editor. The C elegans Research Community. 2006 Available from: < http://www.wormbook.org/chapters/www_chemosensation/chemosensation.html>. - PMC - PubMed
-
- Bargmann CI, Horvitz HR. Chemosensory neurons with overlapping functions direct chemotaxis to multiple chemicals in C. elegans. Neuron. 1991;7:729–742. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
