

Inflammation-induced repression of chromatin bound by the transcription factor Foxp3 in regulatory T cells
- PMID: 24728351
- PMCID: PMC4112080
- DOI: 10.1038/ni.2868
Inflammation-induced repression of chromatin bound by the transcription factor Foxp3 in regulatory T cells
Abstract
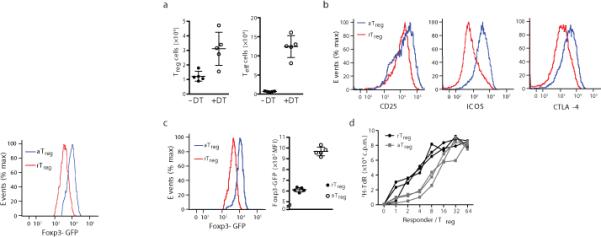
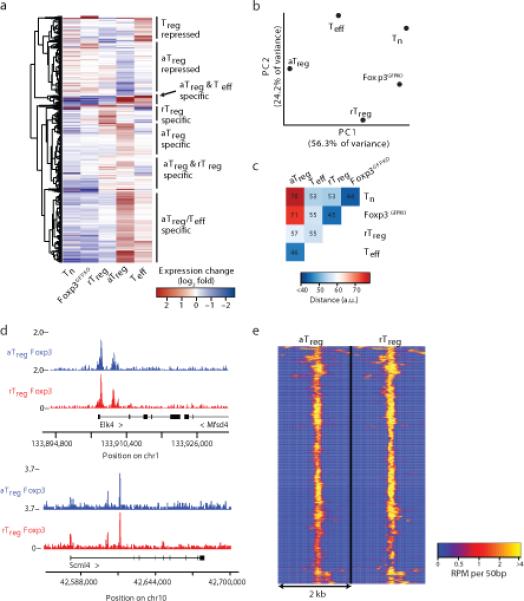
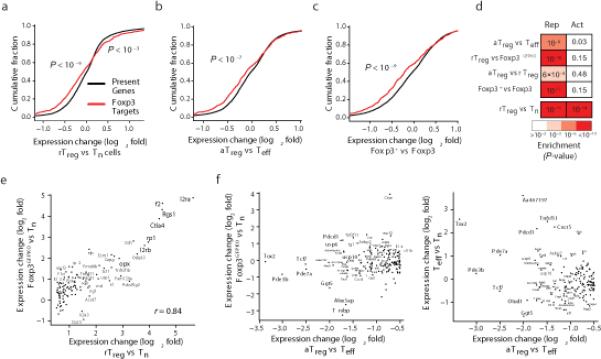
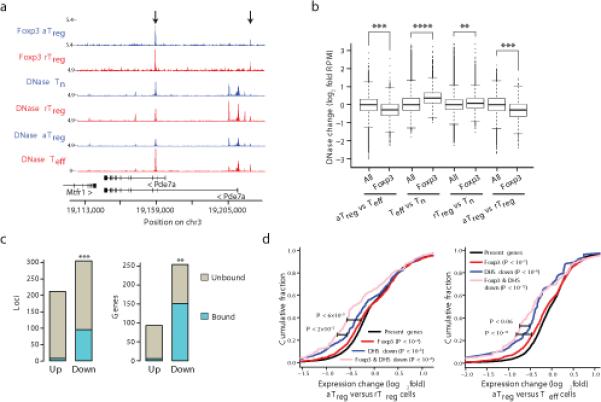
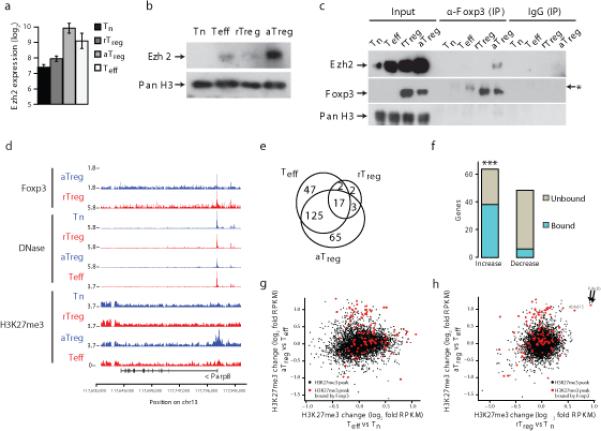
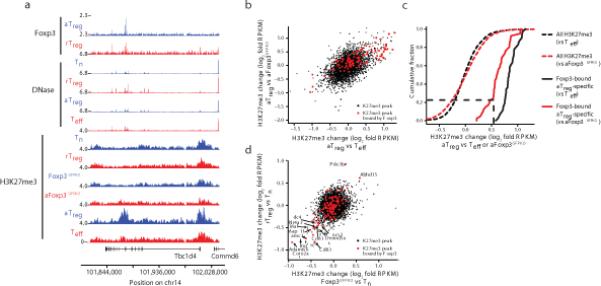
The transcription factor Foxp3 is indispensable for the ability of regulatory T cells (Treg cells) to suppress fatal inflammation. Here we characterized the role of Foxp3 in chromatin remodeling and the regulation of gene expression in actively suppressive Treg cells in an inflammatory setting. Although genome-wide occupancy of regulatory elements in DNA by Foxp3 was similar in resting Treg cells and those activated in vivo, Foxp3-bound enhancer elements in the DNA were poised for repression only in activated Treg cells. Following activation, Foxp3-bound sites showed diminished accessibility of chromatin and selective deposition of histone H3 trimethylated at Lys27 (H3K27me3), which was associated with recruitment of the histone methyltransferase Ezh2 and downregulation of the expression of nearby genes. Thus, Foxp3 poises its targets for repression by facilitating the formation of repressive chromatin in Treg cells upon their activation in response to inflammatory cues.
Figures
References
-
- Ohkura N, Kitagawa Y, Sakaguchi S. Development and maintenance of regulatory T cells. Immunity. 2013;38:414–423. - PubMed
-
- Gavin MA, et al. Foxp3-dependent programme of regulatory T-cell differentiation. Nature. 2007;445:771–775. - PubMed
-
- Lin W, et al. Regulatory T cell development in the absence of functional Foxp3. Nat. Immunol. 2007;8:359–368. - PubMed
-
- Williams LM, Rudensky AY. Maintenance of the Foxp3-dependent developmental program in mature regulatory T cells requires continued expression of Foxp3. Nat. Immunol. 2007;8:277–284. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
