

Site-dependent recruitment of inflammatory cells determines the effective dose of Leishmania major
- PMID: 24733090
- PMCID: PMC4097609
- DOI: 10.1128/IAI.01600-13
Site-dependent recruitment of inflammatory cells determines the effective dose of Leishmania major
Abstract
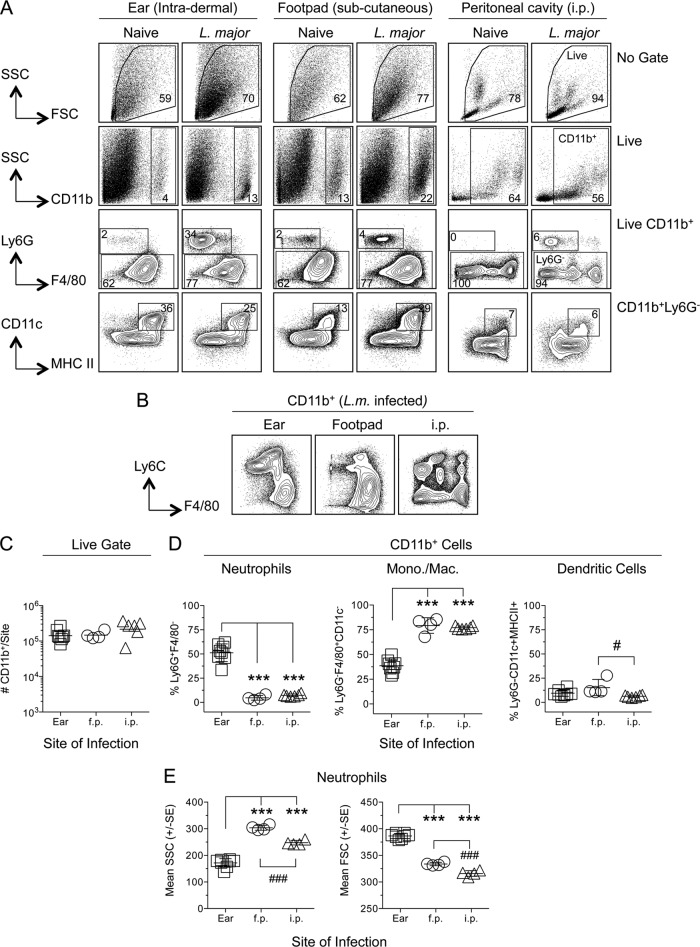
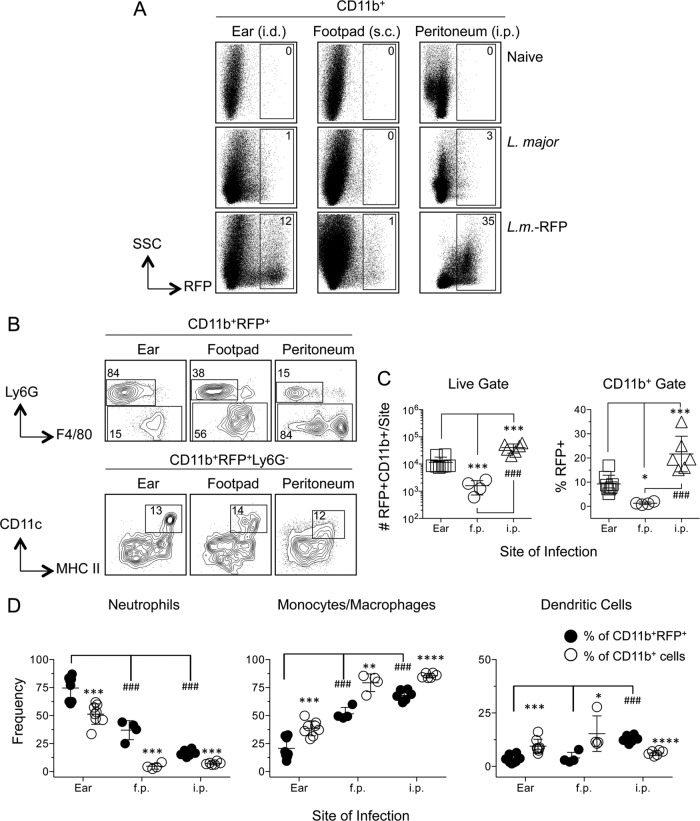
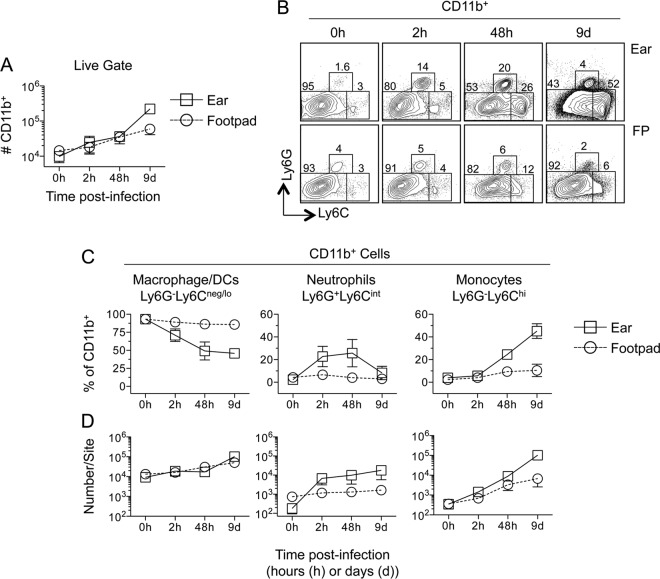
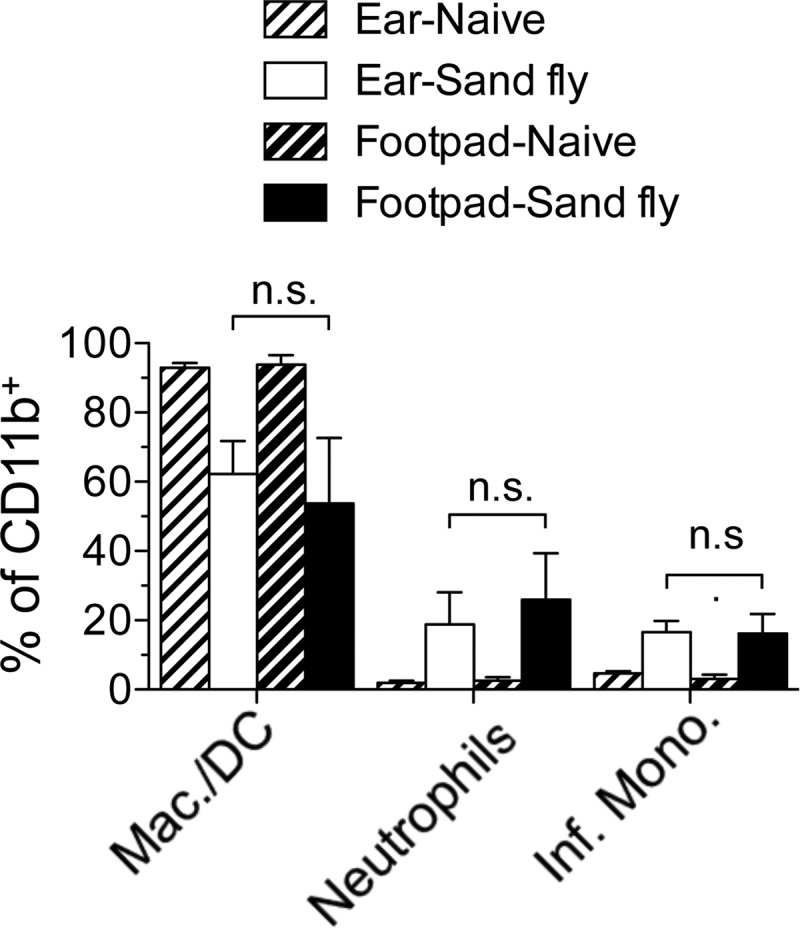
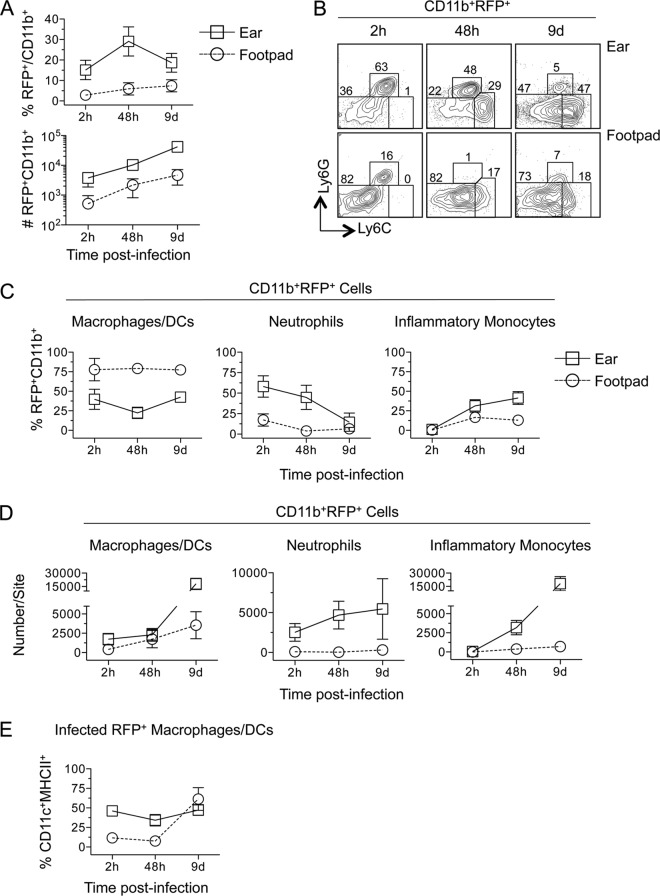
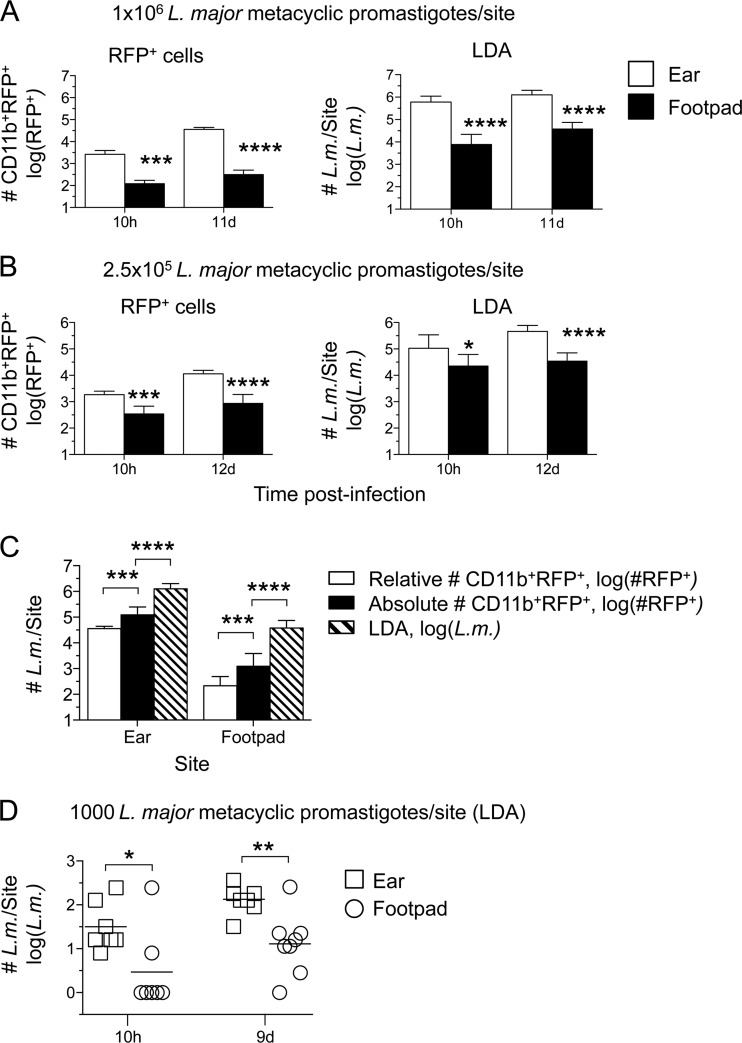
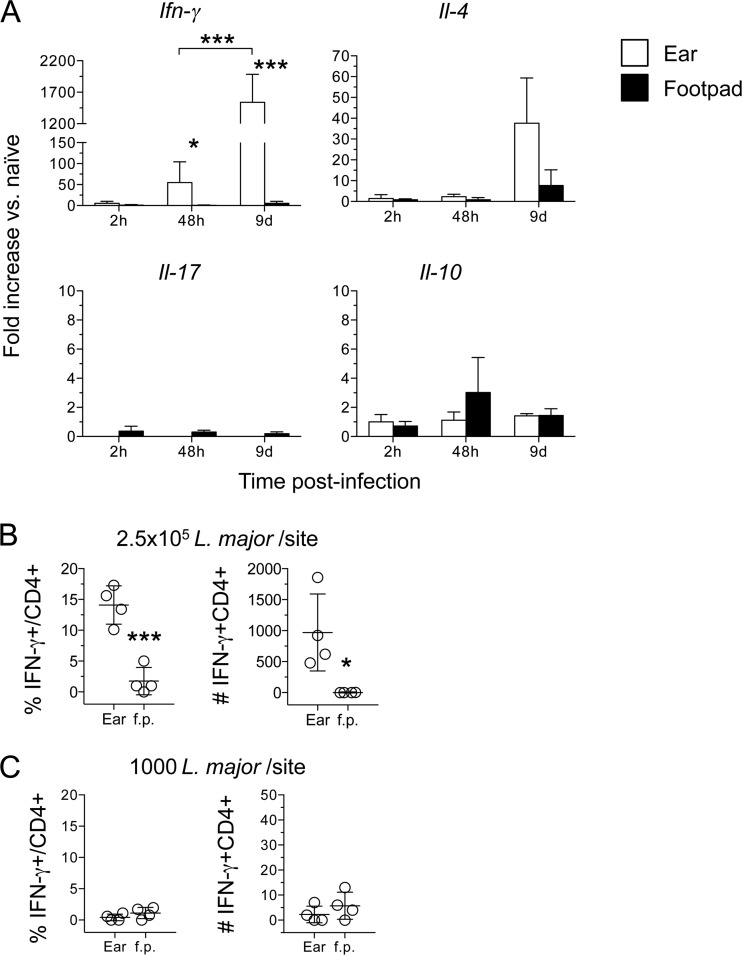
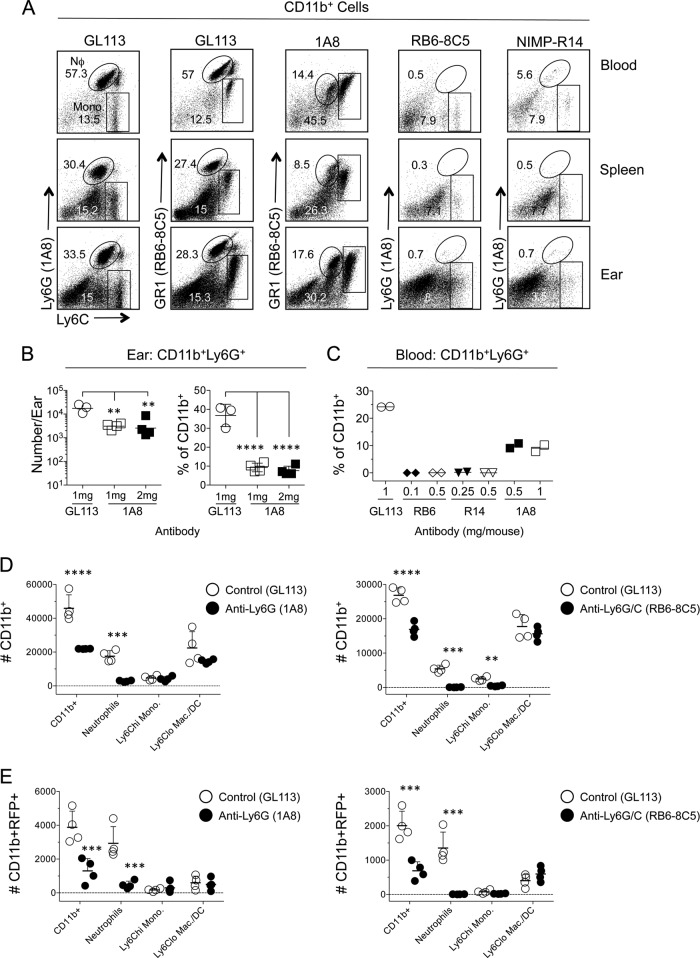
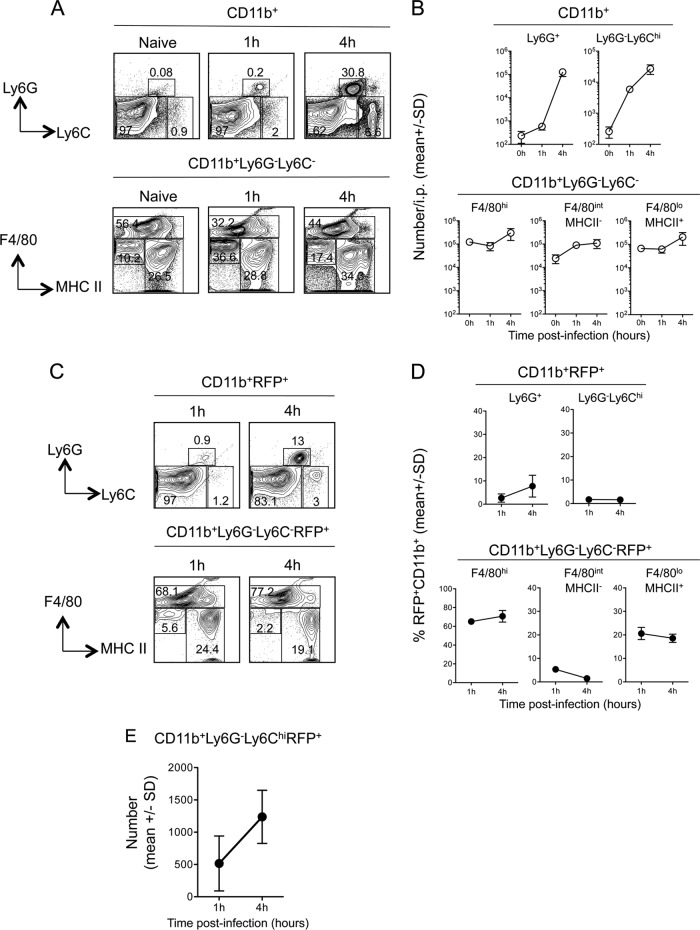
The route of pathogen inoculation by needle has been shown to influence the outcome of infection. Employing needle inoculation of the obligately intracellular parasite Leishmania major, which is transmitted in nature following intradermal (i.d.) deposition of parasites by the bite of an infected sand fly, we identified differences in the preexisting and acute cellular responses in mice following i.d. inoculation of the ear, subcutaneous (s.c.) inoculation of the footpad, or inoculation of the peritoneal cavity (intraperitoneal [i.p.] inoculation). Initiation of infection at different sites was associated with different phagocytic populations. Neutrophils were the dominant infected cells following i.d., but not s.c. or i.p., inoculation. Inoculation of the ear dermis resulted in higher frequencies of total and infected neutrophils than inoculation of the footpad, and these higher frequencies were associated with a 10-fold increase in early parasite loads. Following inoculation of the ear in the absence of neutrophils, parasite phagocytosis by other cell types did not increase, and fewer parasites were able to establish infection. The frequency of infected neutrophils within the total infected CD11b(+) population was higher than the frequency of total neutrophils within the total CD11b(+) population, demonstrating that neutrophils are overrepresented as a proportion of infected cells. Employing i.d. inoculation to model sand fly transmission of parasites has significant consequences for infection outcome relative to that of s.c. or i.p. inoculation, including the phenotype of infected cells and the number of parasites that establish infection. Vector-borne infections initiated in the dermis likely involve adaptations to this unique microenvironment. Bypassing or altering this initial step has significant consequences for infection.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Oliveira DM, Costa MA, Chavez-Fumagalli MA, Valadares DG, Duarte MC, Costa LE, Martins VT, Gomes RF, Melo MN, Soto M, Tavares CA, Coelho EA. 2012. Evaluation of parasitological and immunological parameters of Leishmania chagasi infection in BALB/c mice using different doses and routes of inoculation of parasites. Parasitol. Res. 110:1277–1285. 10.1007/s00436-011-2628-5 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
