

Completely ES cell-derived mice produced by tetraploid complementation using inner cell mass (ICM) deficient blastocysts
- PMID: 24733255
- PMCID: PMC3986396
- DOI: 10.1371/journal.pone.0094730
Completely ES cell-derived mice produced by tetraploid complementation using inner cell mass (ICM) deficient blastocysts
Abstract
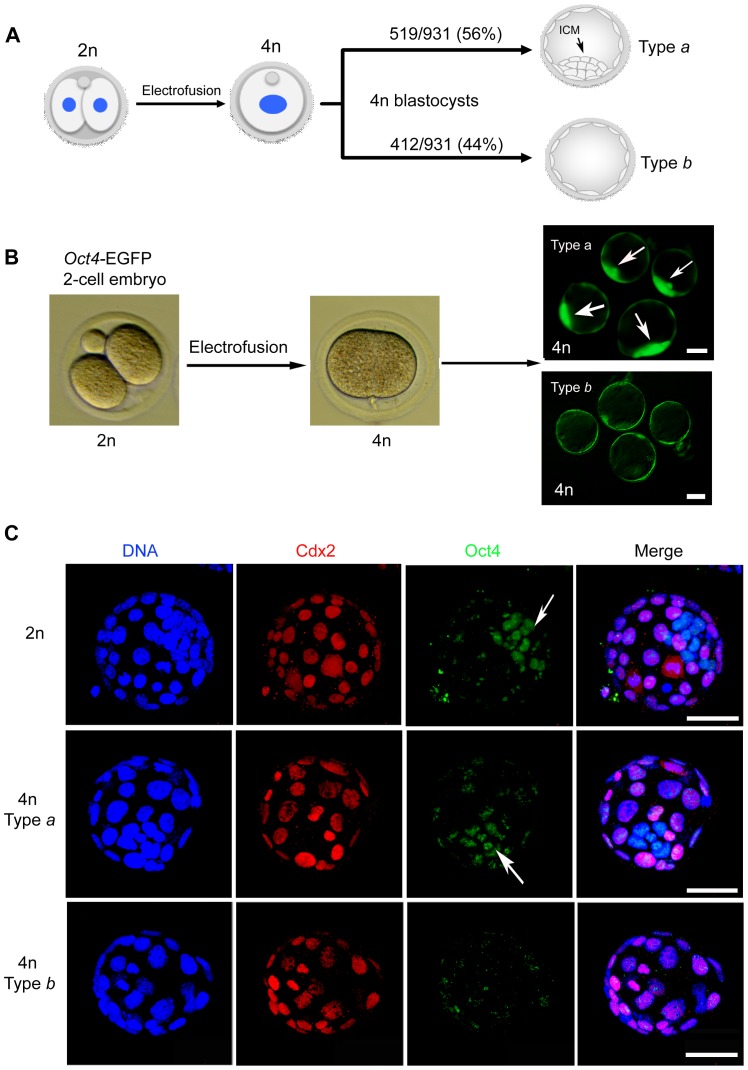
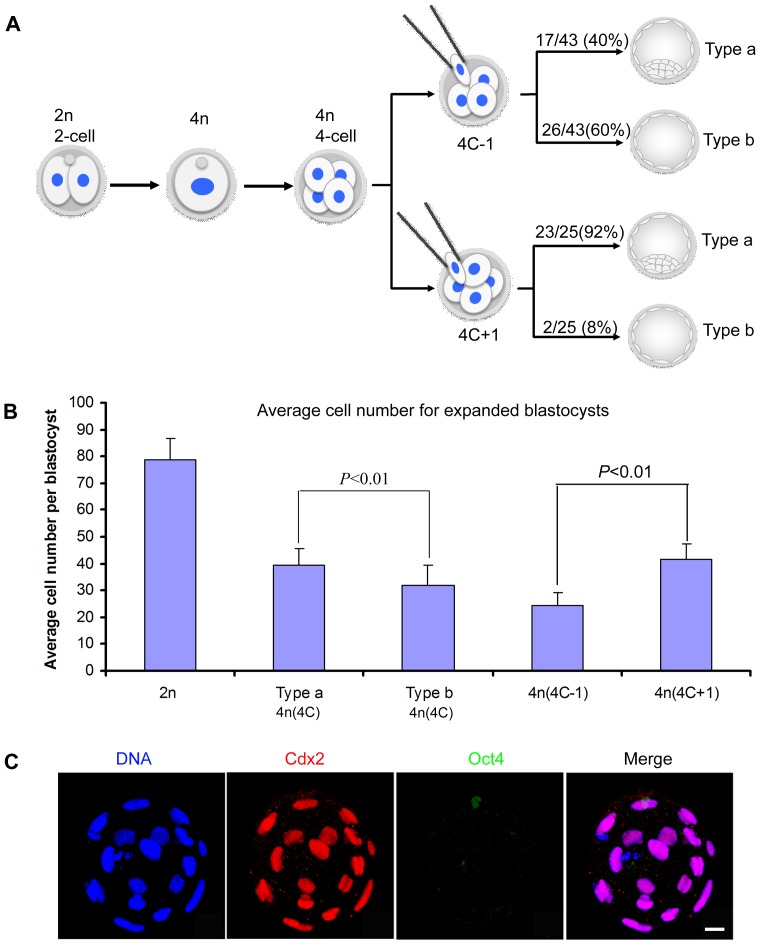
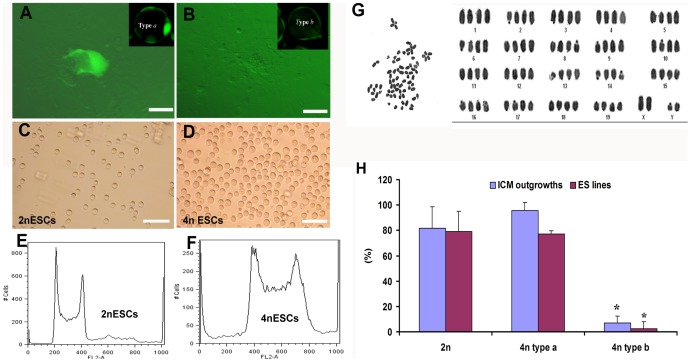
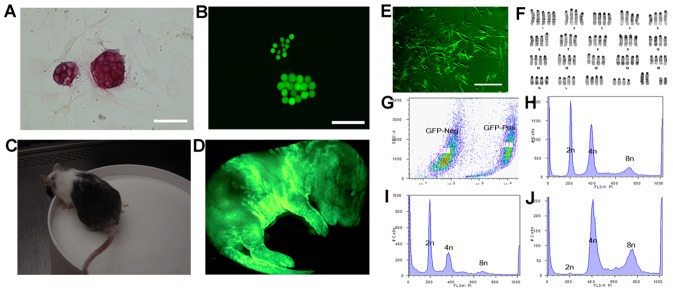
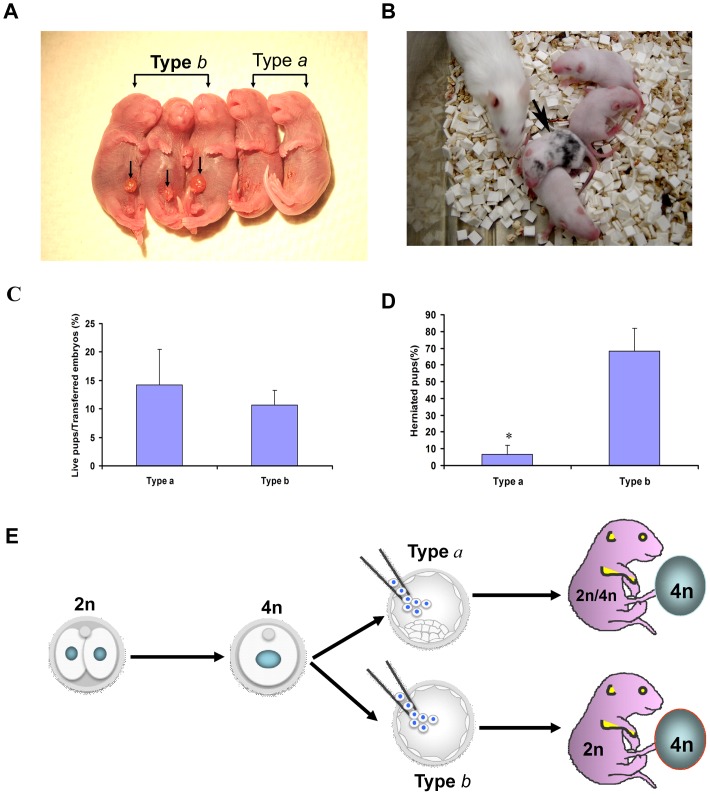
Tetraploid complementation is often used to produce mice from embryonic stem cells (ESCs) by injection of diploid (2n) ESCs into tetraploid (4n) blastocysts (ESC-derived mice). This method has also been adapted to mouse cloning and the derivation of mice from induced pluripotent stem (iPS) cells. However, the underlying mechanism(s) of the tetraploid complementation remains largely unclear. Whether this approach can give rise to completely ES cell-derived mice is an open question, and has not yet been unambiguously proven. Here, we show that mouse tetraploid blastocysts can be classified into two groups, according to the presence or absence of an inner cell mass (ICM). We designate these as type a (presence of ICM at blastocyst stage) or type b (absence of ICM). ESC lines were readily derived from type a blastocysts, suggesting that these embryos retain a pluripotent epiblast compartment; whereas the type b blastocysts possessed very low potential to give rise to ESC lines, suggesting that they had lost the pluripotent epiblast. When the type a blastocysts were used for tetraploid complementation, some of the resulting mice were found to be 2n/4n chimeric; whereas when type b blastocysts were used as hosts, the resulting mice are all completely ES cell-derived, with the newborn pups displaying a high frequency of abdominal hernias. Our results demonstrate that completely ES cell-derived mice can be produced using ICM-deficient 4n blastocysts, and provide evidence that the exclusion of tetraploid cells from the fetus in 2n/4n chimeras can largely be attributed to the formation of ICM-deficient blastocysts.
Conflict of interest statement
Figures
Similar articles
-
Generation and developmental characteristics of porcine tetraploid embryos and tetraploid/diploid chimeric embryos.Genomics Proteomics Bioinformatics. 2013 Oct;11(5):327-33. doi: 10.1016/j.gpb.2013.09.007. Epub 2013 Oct 8. Genomics Proteomics Bioinformatics. 2013. PMID: 24120753 Free PMC article.
-
Fate of tetraploid cells in 4n<-->2n chimeric mouse blastocysts.Mech Dev. 2005 Dec;122(12):1266-81. doi: 10.1016/j.mod.2005.09.001. Epub 2005 Nov 4. Mech Dev. 2005. PMID: 16274964
-
Simultaneous Derivation of Embryonic and Trophoblast Stem Cells from Mouse Blastocysts.Methods Mol Biol. 2020;2117:235-241. doi: 10.1007/978-1-0716-0301-7_14. Methods Mol Biol. 2020. PMID: 31960383
-
The post-inner cell mass intermediate: implications for stem cell biology and assisted reproductive technology.Hum Reprod Update. 2015 Sep-Oct;21(5):616-26. doi: 10.1093/humupd/dmv028. Epub 2015 Jun 18. Hum Reprod Update. 2015. PMID: 26089403 Review.
-
Uncovering the true identity of naïve pluripotent stem cells.Trends Cell Biol. 2013 Sep;23(9):442-8. doi: 10.1016/j.tcb.2013.04.004. Epub 2013 May 17. Trends Cell Biol. 2013. PMID: 23685019 Review.
Cited by
-
Tetraploid Embryonic Stem Cells Maintain Pluripotency and Differentiation Potency into Three Germ Layers.PLoS One. 2015 Jun 19;10(6):e0130585. doi: 10.1371/journal.pone.0130585. eCollection 2015. PLoS One. 2015. PMID: 26091100 Free PMC article.
-
Regulation of Genomic Output and (Pluri)potency in Regeneration.Annu Rev Genet. 2019 Dec 3;53:327-346. doi: 10.1146/annurev-genet-112618-043733. Epub 2019 Sep 10. Annu Rev Genet. 2019. PMID: 31505134 Free PMC article. Review.
-
Dual role of lipids for genome stability and pluripotency facilitates full potency of mouse embryonic stem cells.Protein Cell. 2023 Aug 1;14(8):591-602. doi: 10.1093/procel/pwad008. Protein Cell. 2023. PMID: 37029701 Free PMC article.
-
Altered apoptosis/autophagy and epigenetic modifications cause the impaired postimplantation octaploid embryonic development in mice.Cell Cycle. 2017 Jan 2;16(1):82-90. doi: 10.1080/15384101.2016.1252884. Epub 2016 Nov 10. Cell Cycle. 2017. PMID: 27830977 Free PMC article.
-
Tetraploid embryonic stem cells can contribute to the development of chimeric fetuses and chimeric extraembryonic tissues.Sci Rep. 2017 Jun 8;7(1):3030. doi: 10.1038/s41598-017-02783-0. Sci Rep. 2017. PMID: 28596585 Free PMC article.
References
-
- Eakin GS, Behringer RR (2003) Tetraploid development in the mouse. Dev Dyn 228: 751–766. - PubMed
-
- Henery CC, Kaufman MH (1992) Relationship between cell size and nuclear volume in nucleated red blood cells of developmentally matched diploid and tetraploid mouse embryos. J Exp Zool 261: 472–478. - PubMed
-
- Eakin GS, Hadjantonakis AK, Papaioannou VE, Behringer RR (2005) Developmental potential and behavior of tetraploid cells in the mouse embryo. Dev Biol 288: 150–159. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
