

Pneumocystis jirovecii Rtt109, a novel drug target for Pneumocystis pneumonia in immunosuppressed humans
- PMID: 24733475
- PMCID: PMC4068529
- DOI: 10.1128/AAC.02637-14
Pneumocystis jirovecii Rtt109, a novel drug target for Pneumocystis pneumonia in immunosuppressed humans
Abstract
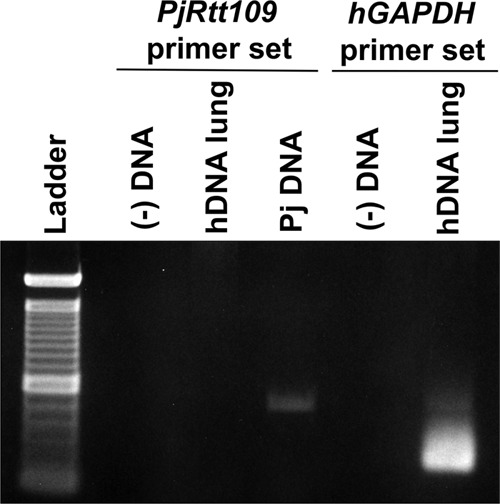
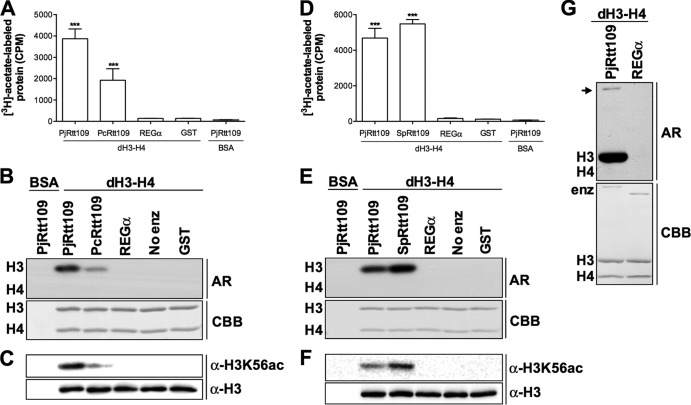
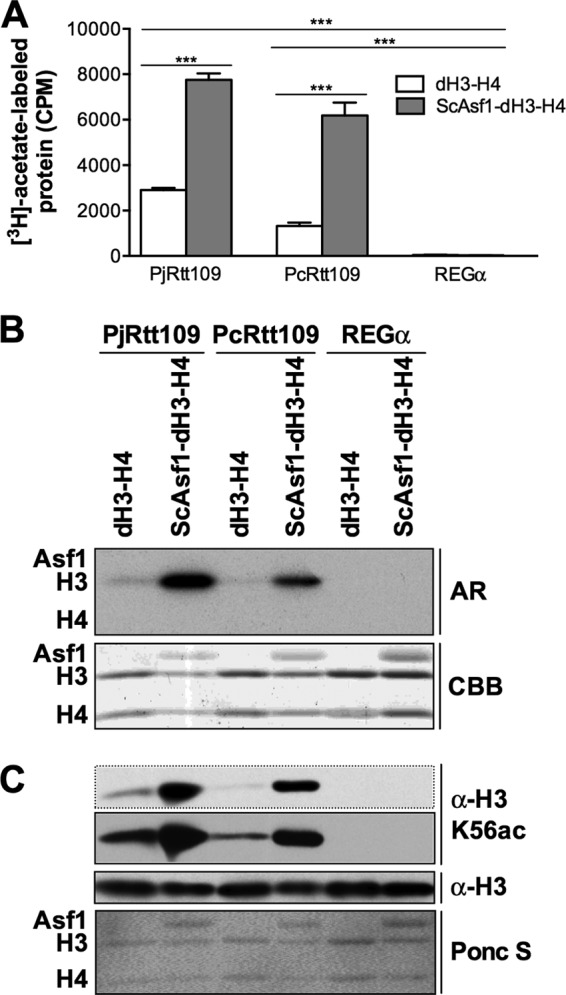
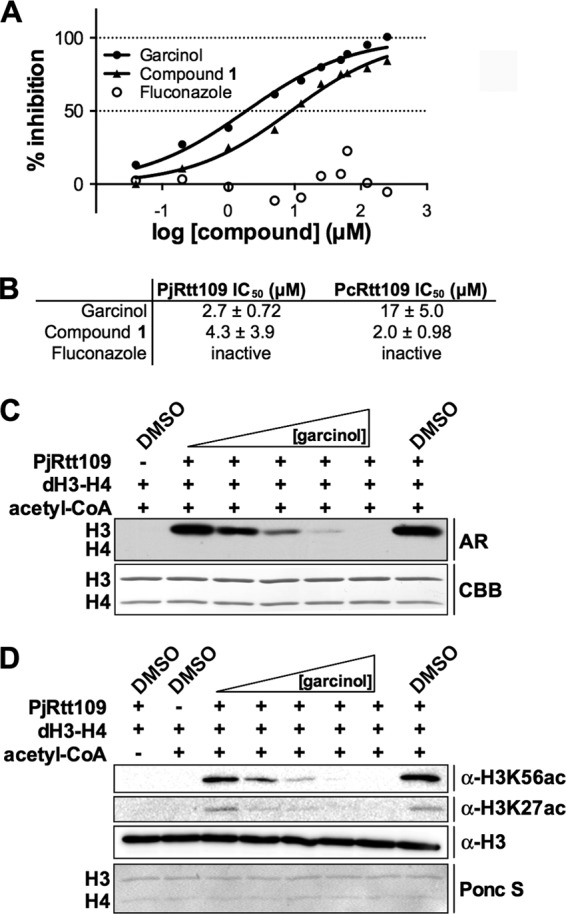
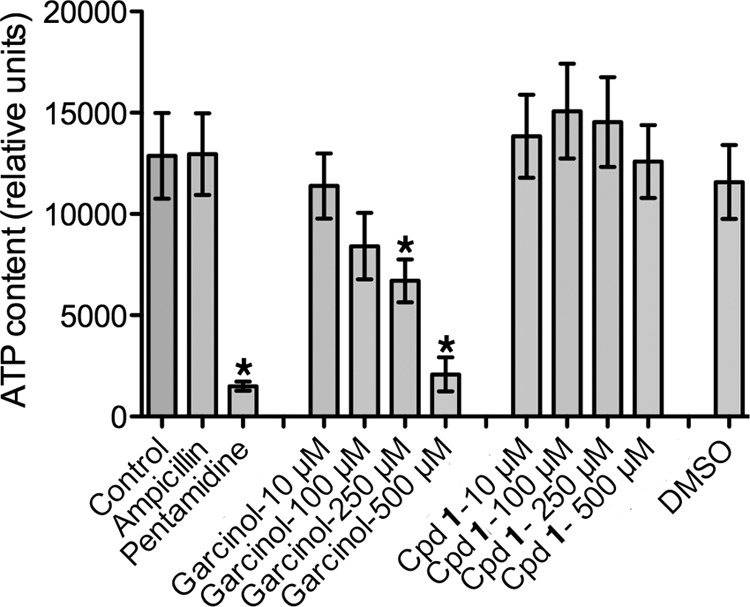
Pneumocystis pneumonia (PcP) is a significant cause of morbidity and mortality in immunocompromised patients. In humans, PcP is caused by the opportunistic fungal species Pneumocystis jirovecii. Progress in Pneumocystis research has been hampered by a lack of viable in vitro culture methods, which limits laboratory access to human-derived organisms for drug testing. Consequently, most basic drug discovery research for P. jirovecii is performed using related surrogate organisms such as Pneumocystis carinii, which is derived from immunosuppressed rodents. While these studies provide useful insights, important questions arise about interspecies variations and the relative utility of identified anti-Pneumocystis agents against human P. jirovecii. Our recent work has identified the histone acetyltransferase (HAT) Rtt109 in P. carinii (i.e., PcRtt109) as a potential therapeutic target for PcP, since Rtt109 HATs are widely conserved in fungi but are absent in humans. To further address the potential utility of this target in human disease, we now demonstrate the presence of a functional Rtt109 orthologue in the clinically relevant fungal pathogen P. jirovecii (i.e., PjRtt109). In a fashion similar to that of Pcrtt109, Pjrtt109 restores H3K56 acetylation and genotoxic resistance in rtt109-null yeast. Recombinant PjRtt109 is an active HAT in vitro, with activity comparable to that of PcRtt109 and yeast Rtt109. PjRtt109 HAT activity is also enhanced by the histone chaperone Asf1 in vitro. PjRtt109 and PcRtt109 showed similar low micromolar sensitivities to two reported small-molecule HAT inhibitors in vitro. Together, these results demonstrate that PjRtt109 is a functional Rtt109 HAT, and they support the development of anti-Pneumocystis agents directed at Rtt109-catalyzed histone acetylation as a novel therapeutic target for human PcP.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures


Similar articles
-
Virtual Screening of Phytochemicals to Novel Target (HAT) Rtt109 in Pneumocystis Jirovecii using Bioinformatics Tools.J Clin Diagn Res. 2016 Mar;10(3):FC05-8. doi: 10.7860/JCDR/2016/16029.7374. Epub 2016 Mar 1. J Clin Diagn Res. 2016. PMID: 27134887 Free PMC article.
-
Pneumocystis carinii expresses an active Rtt109 histone acetyltransferase.Am J Respir Cell Mol Biol. 2011 Jun;44(6):768-76. doi: 10.1165/rcmb.2009-0443OC. Epub 2010 Jul 23. Am J Respir Cell Mol Biol. 2011. PMID: 20656950 Free PMC article.
-
Characterization of the Pneumocystis carinii histone acetyltransferase chaperone proteins PcAsf1 and PcVps75.Infect Immun. 2013 Jun;81(6):2268-75. doi: 10.1128/IAI.01173-12. Epub 2013 Apr 8. Infect Immun. 2013. PMID: 23569117 Free PMC article.
-
Recent Advances in the Diagnosis of Pneumocystis Pneumonia.Med Mycol J. 2016;57(4):E111-E116. doi: 10.3314/mmj.16-00019. Med Mycol J. 2016. PMID: 27904052 Review.
-
Pulmonary co-infections by Pneumocystis jirovecii and Aspergillus fumigatus in non-HIV patients: A report of two cases and literature review.Mycoses. 2017 Oct;60(10):626-633. doi: 10.1111/myc.12642. Epub 2017 Jun 28. Mycoses. 2017. PMID: 28660636 Review.
Cited by
-
Stoichiometry of Rtt109 complexes with Vps75 and histones H3-H4.Life Sci Alliance. 2020 Sep 10;3(11):e202000771. doi: 10.26508/lsa.202000771. Print 2020 Nov. Life Sci Alliance. 2020. PMID: 32913112 Free PMC article.
-
Characterization of N-Acetylglucosamine Biosynthesis in Pneumocystis species. A New Potential Target for Therapy.Am J Respir Cell Mol Biol. 2017 Feb;56(2):213-222. doi: 10.1165/rcmb.2016-0155OC. Am J Respir Cell Mol Biol. 2017. PMID: 27632412 Free PMC article.
-
Virtual Screening of Phytochemicals to Novel Target (HAT) Rtt109 in Pneumocystis Jirovecii using Bioinformatics Tools.J Clin Diagn Res. 2016 Mar;10(3):FC05-8. doi: 10.7860/JCDR/2016/16029.7374. Epub 2016 Mar 1. J Clin Diagn Res. 2016. PMID: 27134887 Free PMC article.
-
Post-HTS case report and structural alert: Promiscuous 4-aroyl-1,5-disubstituted-3-hydroxy-2H-pyrrol-2-one actives verified by ALARM NMR.Bioorg Med Chem Lett. 2015 Nov 1;25(21):4740-4752. doi: 10.1016/j.bmcl.2015.08.020. Epub 2015 Aug 10. Bioorg Med Chem Lett. 2015. PMID: 26318992 Free PMC article.
References
-
- Huang L, Cattamanchi A, Davis JL, den Boon S, Kovacs J, Meshnick S, Miller RF, Walzer PD, Worodria W, Masur H, International HIV-Associated Opportunistic Pneumonias (IHOP) Study, Lung HIV Study 2011. HIV-associated Pneumocystis pneumonia. Proc. Am. Thorac. Soc. 8:294–300. 10.1513/pats.201009-062WR - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
